

Immune Mechanisms in Inflammatory Anemia
- PMID: 36750316
- PMCID: PMC10367595
- DOI: 10.1146/annurev-immunol-101320-125839
Immune Mechanisms in Inflammatory Anemia
Abstract
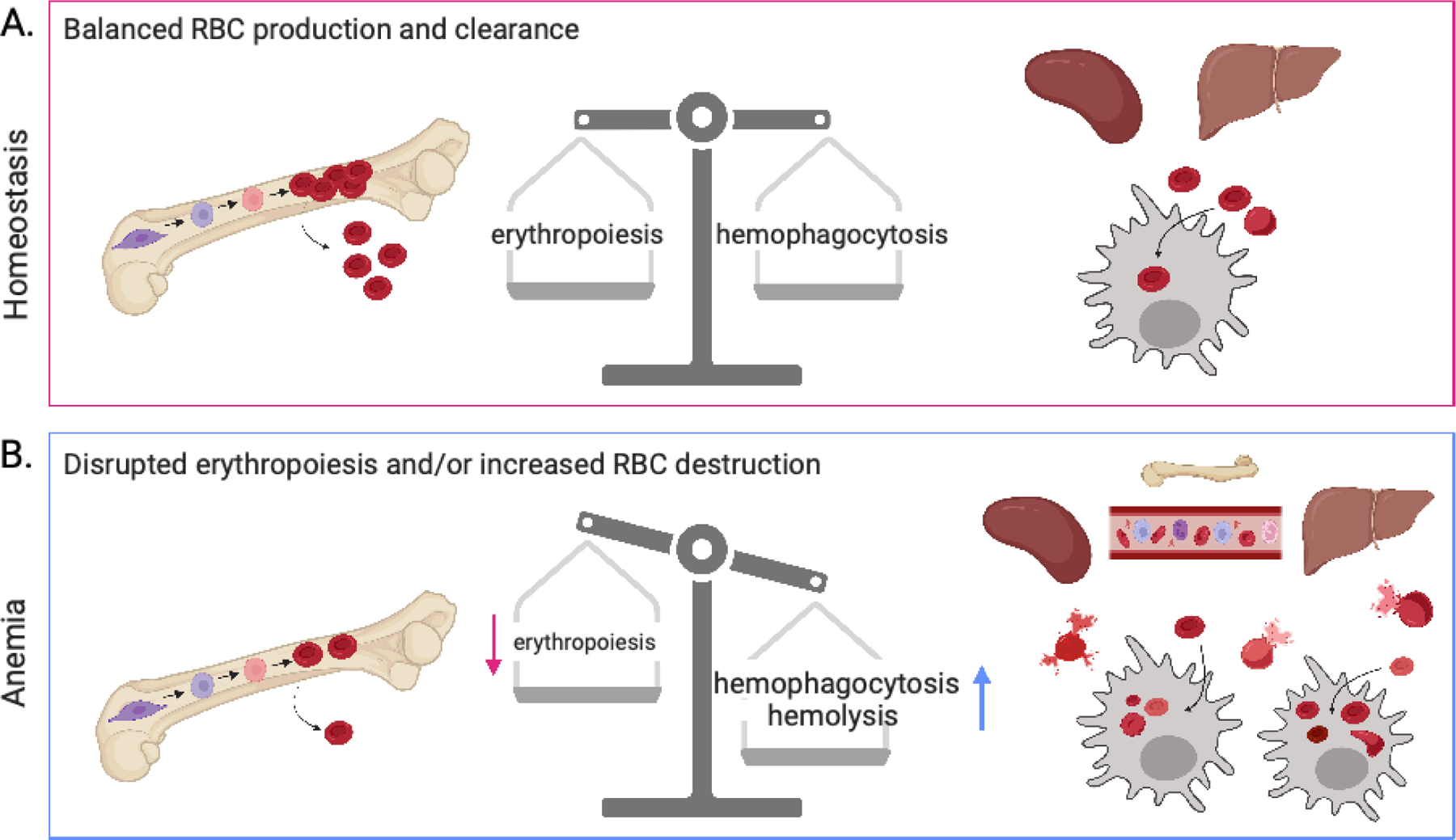
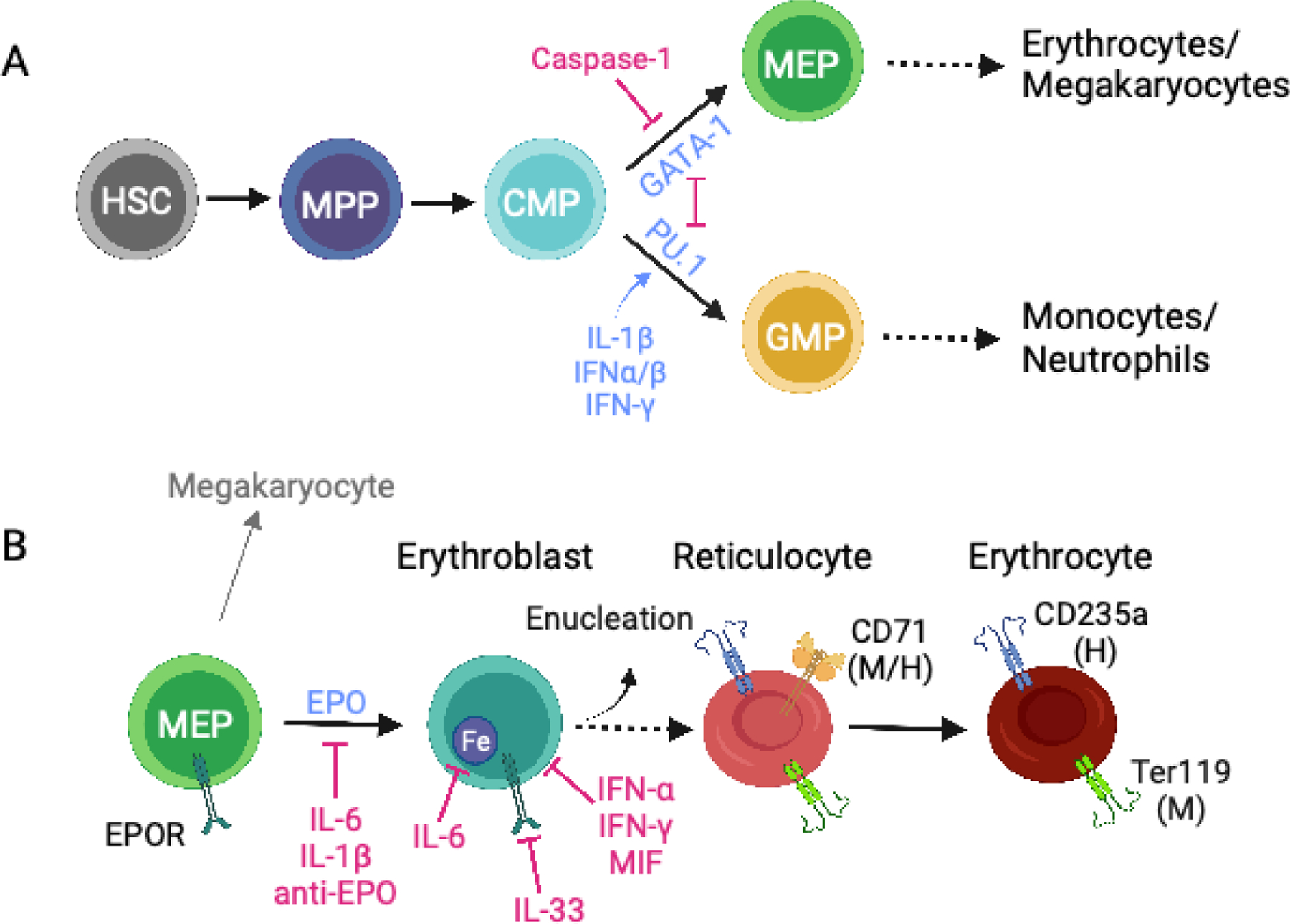
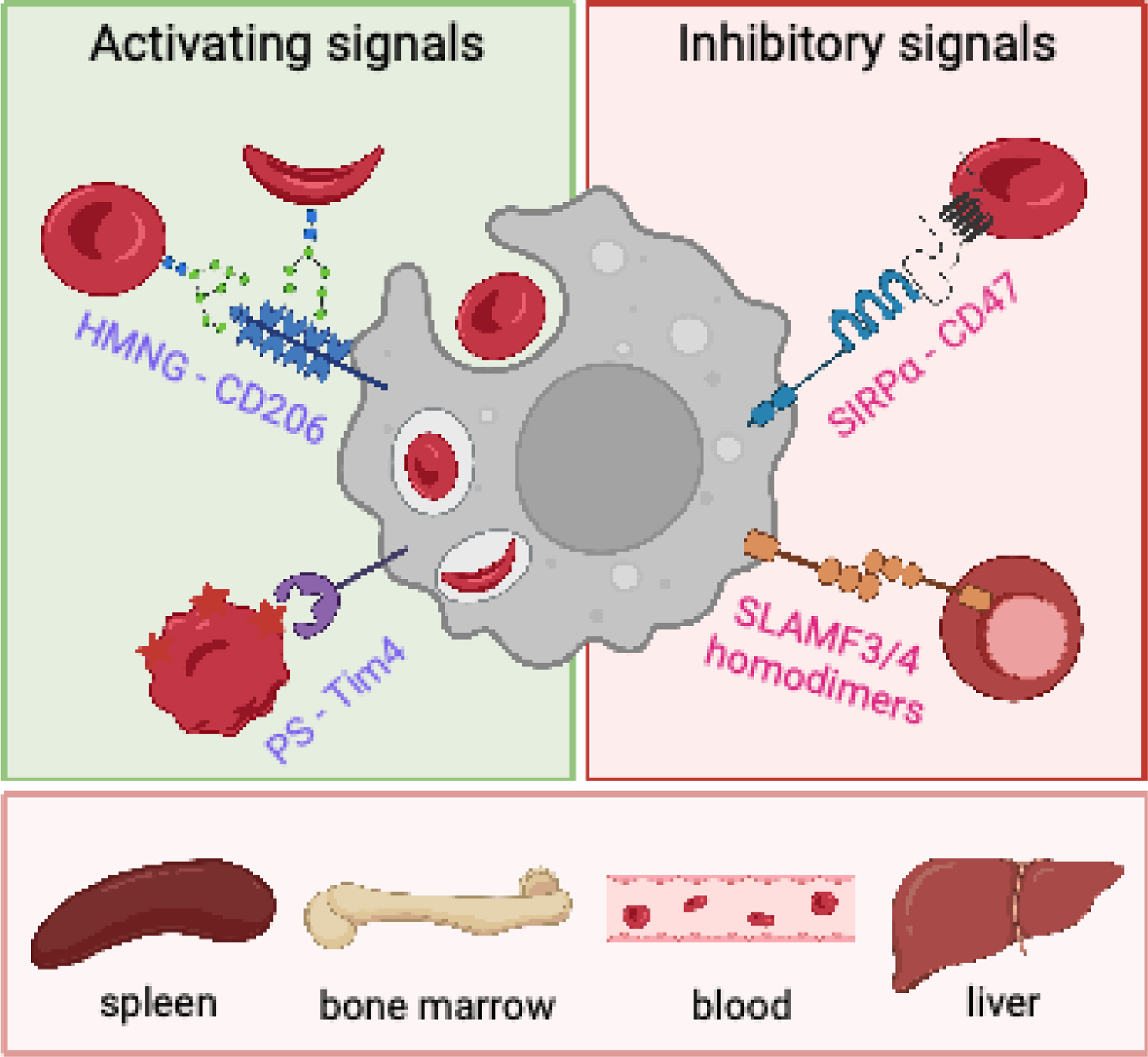
Maintaining the correct number of healthy red blood cells (RBCs) is critical for proper oxygenation of tissues throughout the body. Therefore, RBC homeostasis is a tightly controlled balance between RBC production and RBC clearance, through the processes of erythropoiesis and macrophage hemophagocytosis, respectively. However, during the inflammation associated with infectious, autoimmune, or inflammatory diseases this homeostatic process is often dysregulated, leading to acute or chronic anemia. In each disease setting, multiple mechanisms typically contribute to the development of inflammatory anemia, impinging on both sides of the RBC production and RBC clearance equation. These mechanisms include both direct and indirect effects of inflammatory cytokines and innate sensing. Here, we focus on common innate and adaptive immune mechanisms that contribute to inflammatory anemias using examples from several diseases, including hemophagocytic lymphohistiocytosis/macrophage activation syndrome, severe malarial anemia during Plasmodium infection, and systemic lupus erythematosus, among others.
Keywords: anemia; hemophagocyte; hemophagocytic lymphohistiocytosis; inflammation; malaria; sickle cell anemia.
Figures
References
-
- Nairz M, Weiss G. 2020. Iron in infection and immunity. Mol. Aspects Med 75:100864. - PubMed
-
- Schultze JL, Mass E, Schlitzer A. 2019. Emerging principles in myelopoiesis at homeostasis and during infection and inflammation. Immunity 50(2):288–301 - PubMed
-
- Doulatov S, Notta F, Laurenti E, Dick JE. 2012. Hematopoiesis: a human perspective. Cell Stem Cell 10(2):120–36 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
