

Myristic acid as a checkpoint to regulate STING-dependent autophagy and interferon responses by promoting N-myristoylation
- PMID: 36750575
- PMCID: PMC9905541
- DOI: 10.1038/s41467-023-36332-3
Myristic acid as a checkpoint to regulate STING-dependent autophagy and interferon responses by promoting N-myristoylation
Abstract
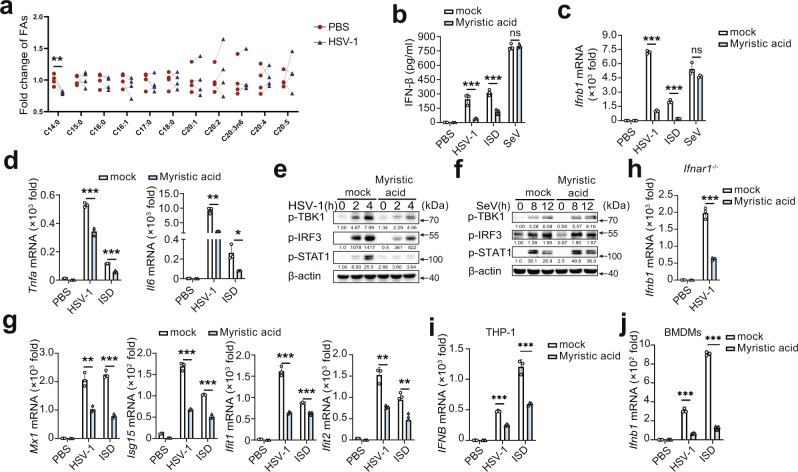
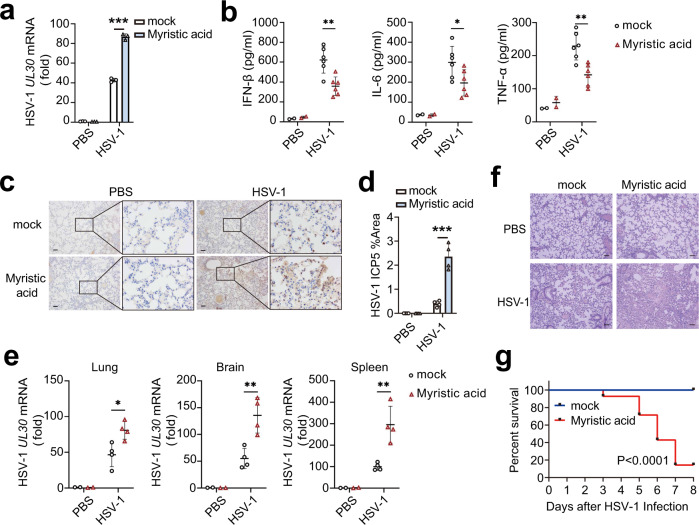
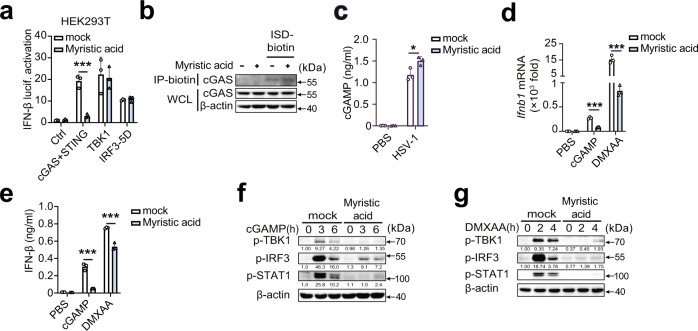
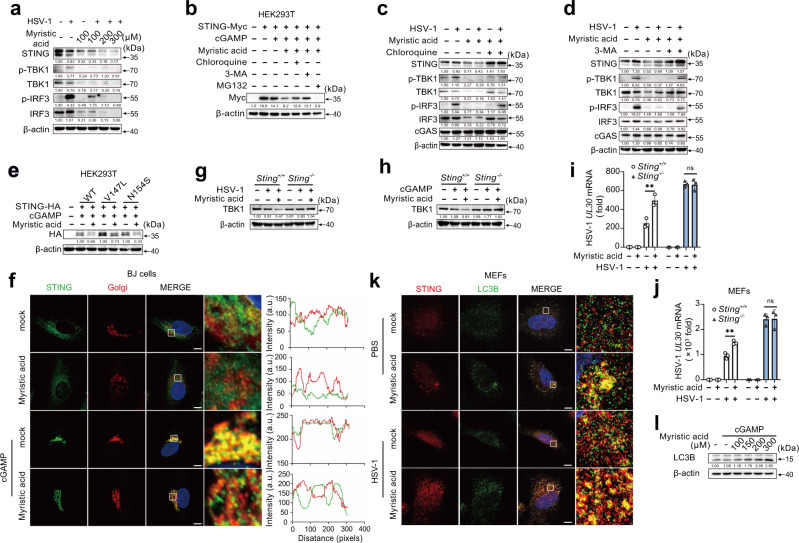
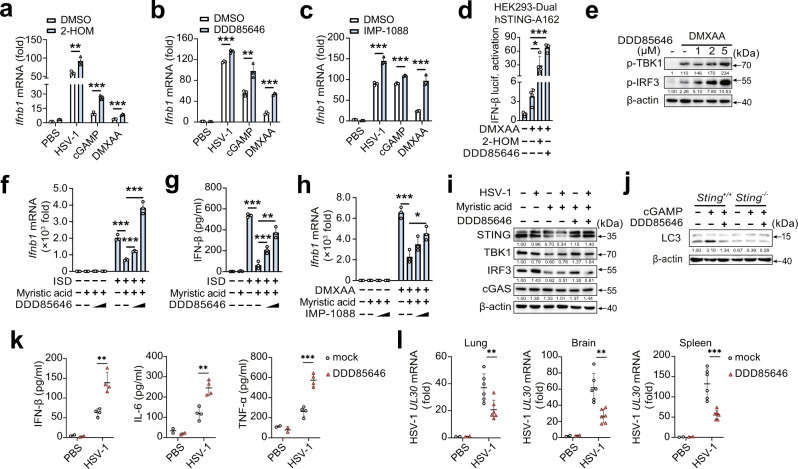
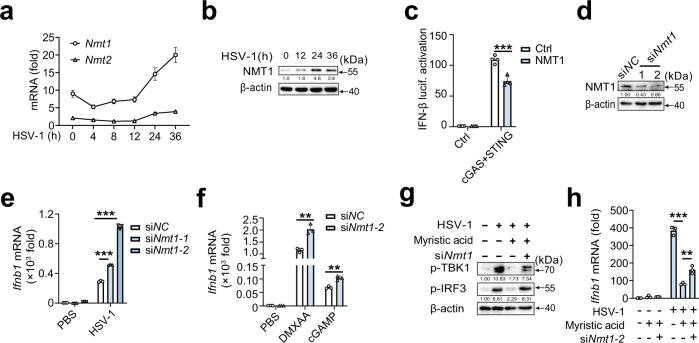
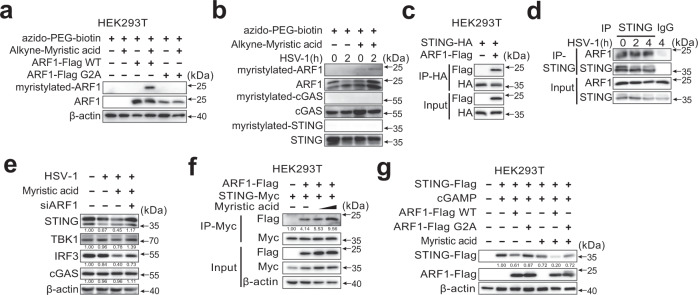
Stimulator of interferon gene (STING)-triggered autophagy is crucial for the host to eliminate invading pathogens and serves as a self-limiting mechanism of STING-induced interferon (IFN) responses. Thus, the mechanisms that ensure the beneficial effects of STING activation are of particular importance. Herein, we show that myristic acid, a type of long-chain saturated fatty acid (SFA), specifically attenuates cGAS-STING-induced IFN responses in macrophages, while enhancing STING-dependent autophagy. Myristic acid inhibits HSV-1 infection-induced innate antiviral immune responses and promotes HSV-1 replication in mice in vivo. Mechanistically, myristic acid enhances N-myristoylation of ARF1, a master regulator that controls STING membrane trafficking. Consequently, myristic acid facilitates STING activation-triggered autophagy degradation of the STING complex. Thus, our work identifies myristic acid as a metabolic checkpoint that contributes to immune homeostasis by balancing STING-dependent autophagy and IFN responses. This suggests that myristic acid and N-myristoylation are promising targets for the treatment of diseases caused by aberrant STING activation.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Chen Q, Sun L, Chen ZJ. Regulation and function of the cGAS-STING pathway of cytosolic DNA sensing. Nat. Immunol. 2016;17:1142–1149. - PubMed
-
- Hopfner KP, Hornung V. Molecular mechanisms and cellular functions of cGAS-STING signalling. Nat. Rev. Mol. Cell Biol. 2020;21:501–521. - PubMed
-
- Motwani M, Pesiridis S, Fitzgerald KA. DNA sensing by the cGAS-STING pathway in health and disease. Nat. Rev. Genet. 2019;20:657–674. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
