

How our understanding of memory replay evolves
- PMID: 36752404
- PMCID: PMC9988534
- DOI: 10.1152/jn.00454.2022
How our understanding of memory replay evolves
Abstract
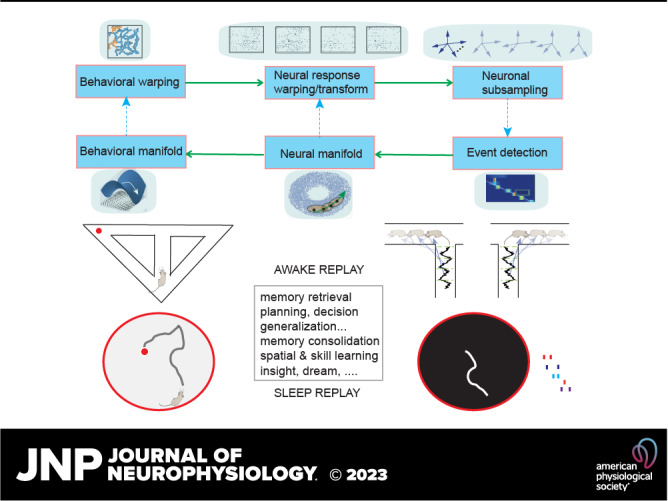
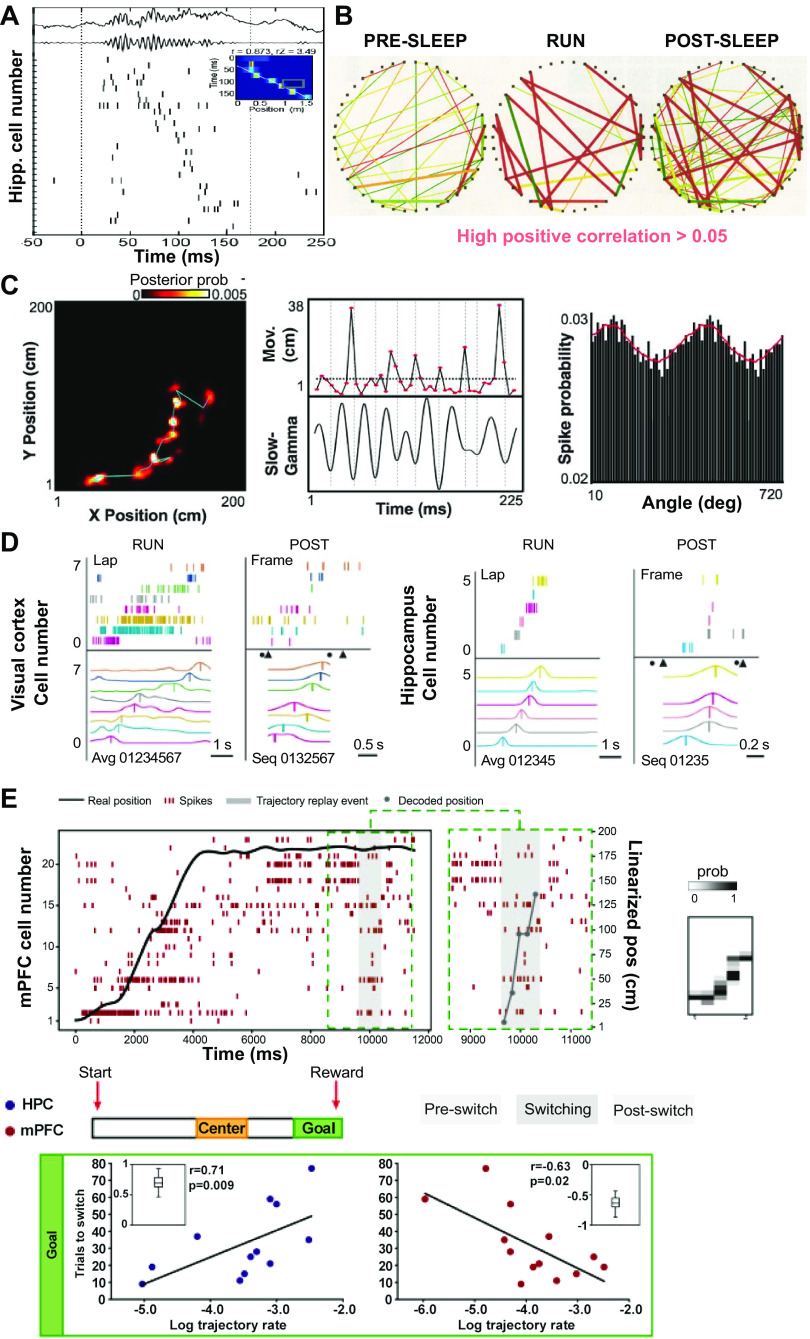
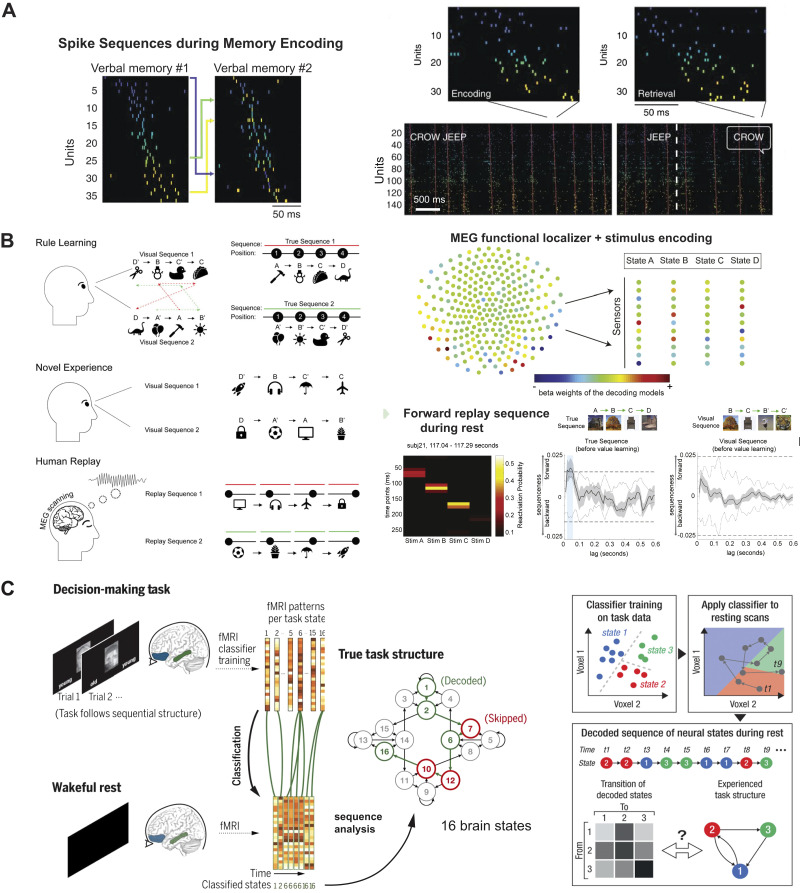
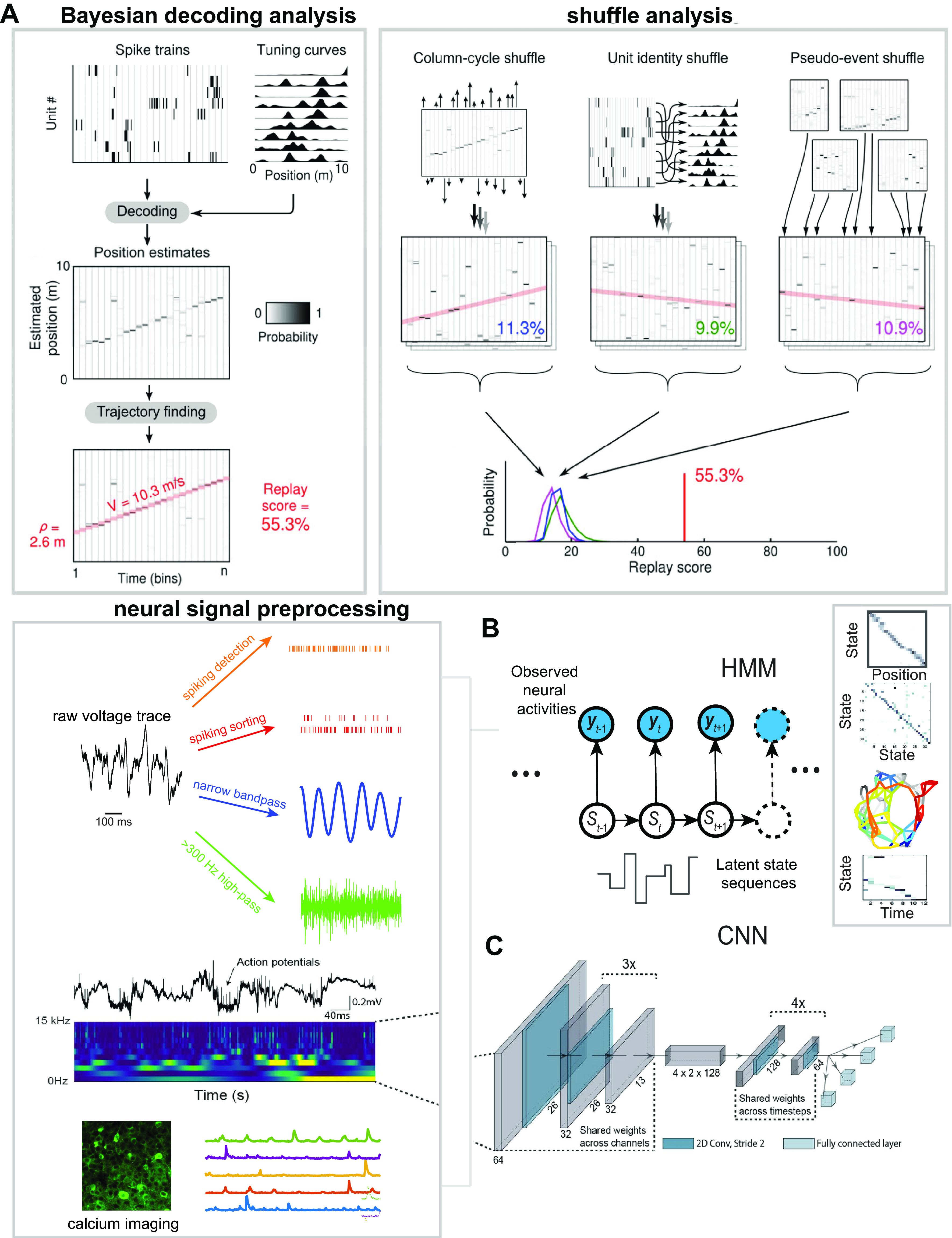
Memory reactivations and replay, widely reported in the hippocampus and cortex across species, have been implicated in memory consolidation, planning, and spatial and skill learning. Technological advances in electrophysiology, calcium imaging, and human neuroimaging techniques have enabled neuroscientists to measure large-scale neural activity with increasing spatiotemporal resolution and have provided opportunities for developing robust analytic methods to identify memory replay. In this article, we first review a large body of historically important and representative memory replay studies from the animal and human literature. We then discuss our current understanding of memory replay functions in learning, planning, and memory consolidation and further discuss the progress in computational modeling that has contributed to these improvements. Next, we review past and present analytic methods for replay analyses and discuss their limitations and challenges. Finally, looking ahead, we discuss some promising analytic methods for detecting nonstereotypical, behaviorally nondecodable structures from large-scale neural recordings. We argue that seamless integration of multisite recordings, real-time replay decoding, and closed-loop manipulation experiments will be essential for delineating the role of memory replay in a wide range of cognitive and motor functions.
Keywords: memory replay; population decoding; representational similarity analysis; sleep replay; temporally delayed linear modeling.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the authors.
Figures

Similar articles
-
A neural network account of memory replay and knowledge consolidation.Cereb Cortex. 2022 Dec 15;33(1):83-95. doi: 10.1093/cercor/bhac054. Cereb Cortex. 2022. PMID: 35213689 Free PMC article.
-
Learned Motor Patterns Are Replayed in Human Motor Cortex during Sleep.J Neurosci. 2022 Jun 22;42(25):5007-5020. doi: 10.1523/JNEUROSCI.2074-21.2022. Epub 2022 May 19. J Neurosci. 2022. PMID: 35589391 Free PMC article.
-
Hippocampal Reactivation Extends for Several Hours Following Novel Experience.J Neurosci. 2019 Jan 30;39(5):866-875. doi: 10.1523/JNEUROSCI.1950-18.2018. Epub 2018 Dec 10. J Neurosci. 2019. PMID: 30530857 Free PMC article.
-
The content of hippocampal "replay".Hippocampus. 2020 Jan;30(1):6-18. doi: 10.1002/hipo.22824. Epub 2018 Jan 10. Hippocampus. 2020. PMID: 29266510 Free PMC article. Review.
-
Replay Comes of Age.Annu Rev Neurosci. 2017 Jul 25;40:581-602. doi: 10.1146/annurev-neuro-072116-031538. Annu Rev Neurosci. 2017. PMID: 28772098 Review.
Cited by
-
Song-like activation of syringeal and respiratory muscles during sleep in canaries.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2025 Mar;211(2):135-148. doi: 10.1007/s00359-024-01720-7. Epub 2024 Oct 15. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2025. PMID: 39404890
-
Respiration modulates sleep oscillations and memory reactivation in humans.Nat Commun. 2023 Dec 18;14(1):8351. doi: 10.1038/s41467-023-43450-5. Nat Commun. 2023. PMID: 38110418 Free PMC article.
-
Reactivation strength during cued recall is modulated by graph distance within cognitive maps.Elife. 2024 May 29;12:RP93357. doi: 10.7554/eLife.93357. Elife. 2024. PMID: 38810249 Free PMC article.
-
Learning of object-in-context sequences in freely-moving macaques.bioRxiv [Preprint]. 2023 Dec 12:2023.12.11.571113. doi: 10.1101/2023.12.11.571113. bioRxiv. 2023. PMID: 38168449 Free PMC article. Preprint.
-
Replay and representation dynamics in the hippocampus of freely flying bats.Nature. 2025 Jul 9. doi: 10.1038/s41586-025-09341-z. Online ahead of print. Nature. 2025. PMID: 40633570
References
-
- Ryle G. The Concept of Mind. Chicago, IL: University of Chicago Press, 1949.
-
- Chung JE, Joo HR, Fan JL, Liu DF, Barnett AH, Chen S, Geaghan-Breiner C, Karlsson MP, Karlsson M, Lee KY, Liang H, Magland JF, Pebbles JA, Tooker AC, Greengard LF, Tolosa VM, Frank LM. High-density, long-lasting, and multi-region electrophysiological recordings using polymer electrode arrays. Neuron, 101: 21–31.e5, 2019. doi:10.1016/j.neuron.2018.11.002. - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous