

Motor processivity and speed determine structure and dynamics of microtubule-motor assemblies
- PMID: 36752605
- PMCID: PMC10014072
- DOI: 10.7554/eLife.79402
Motor processivity and speed determine structure and dynamics of microtubule-motor assemblies
Abstract
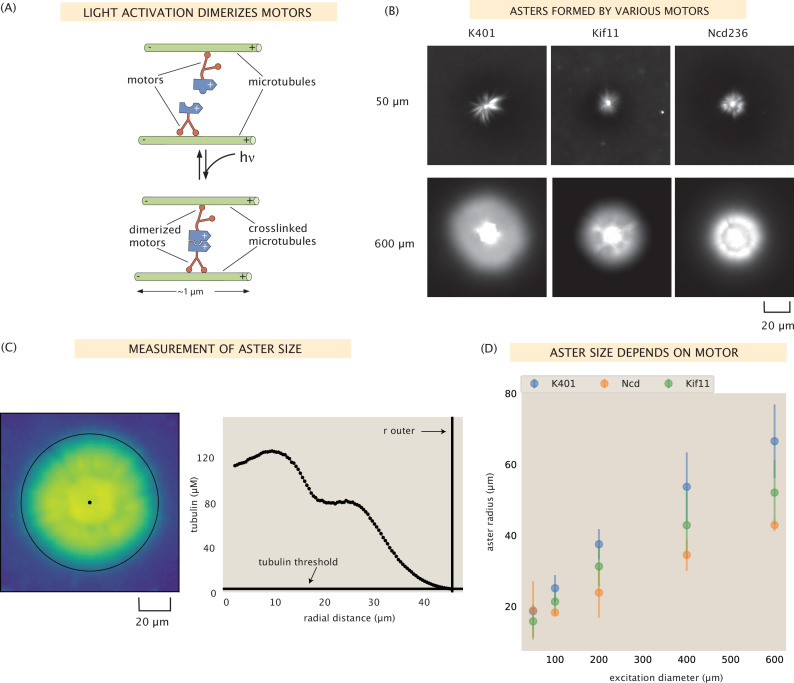
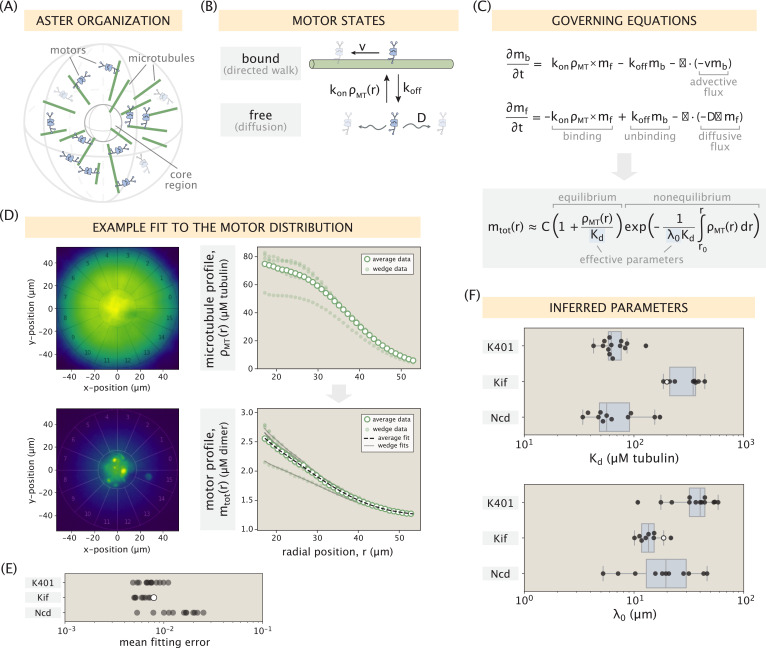
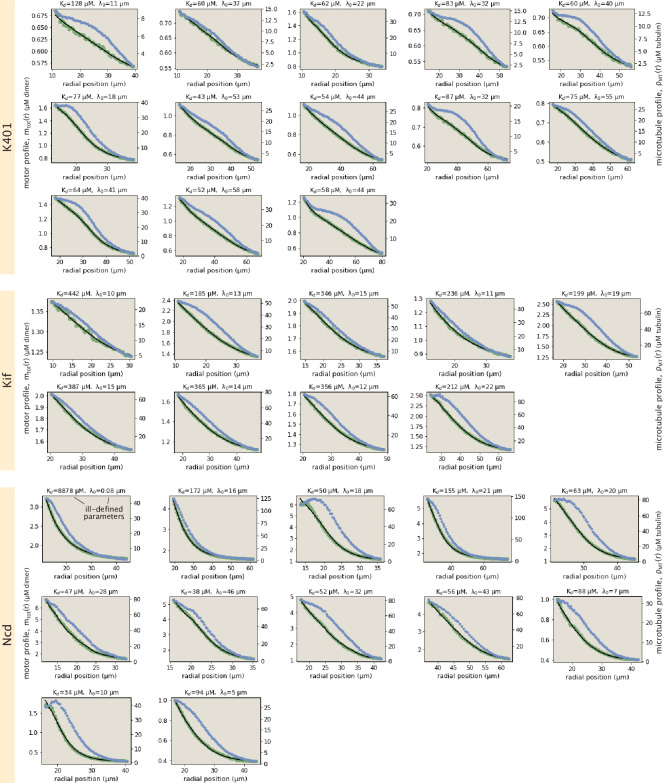
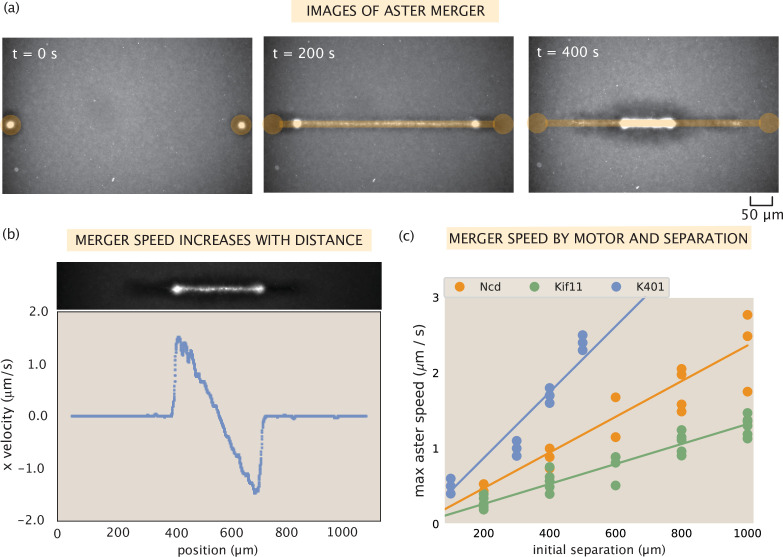
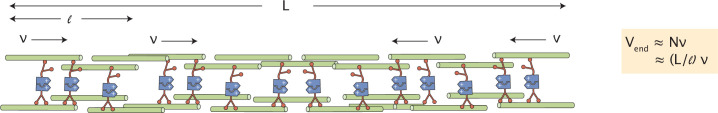
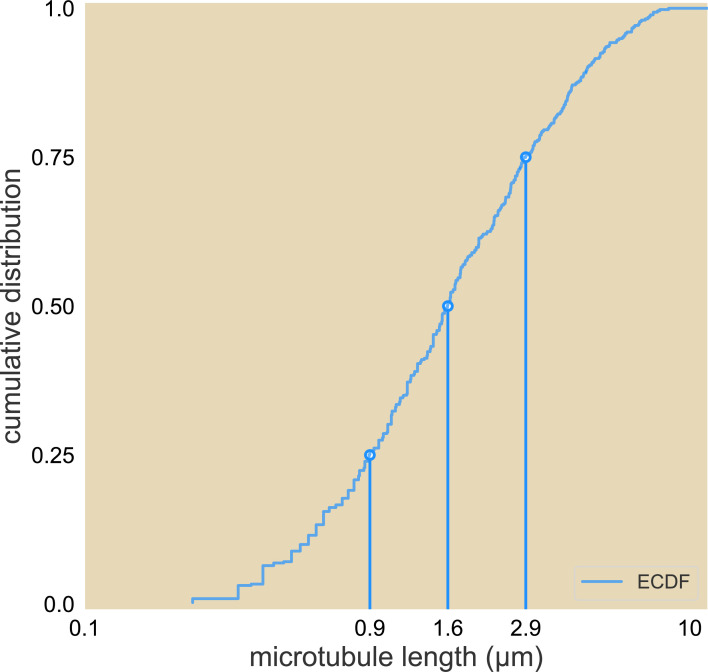
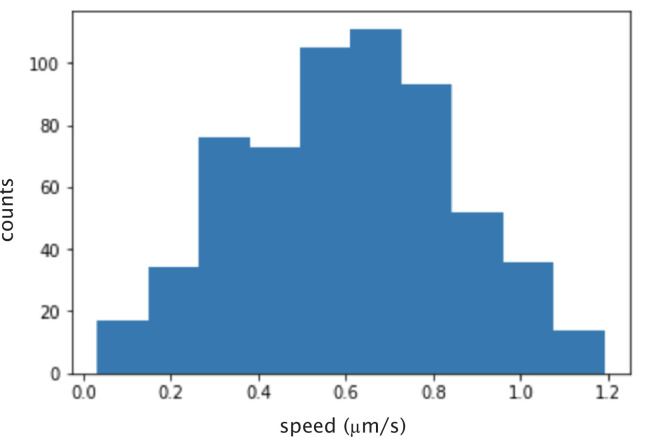
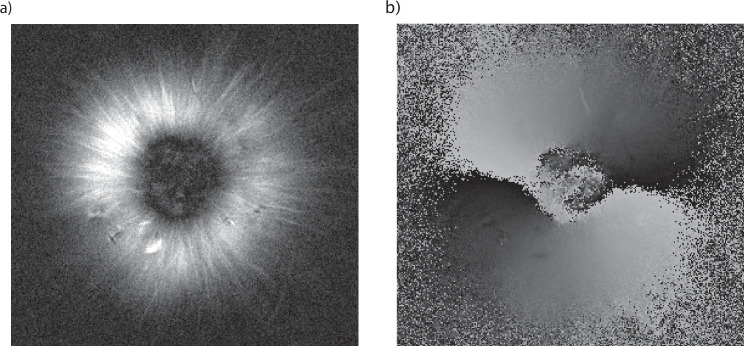
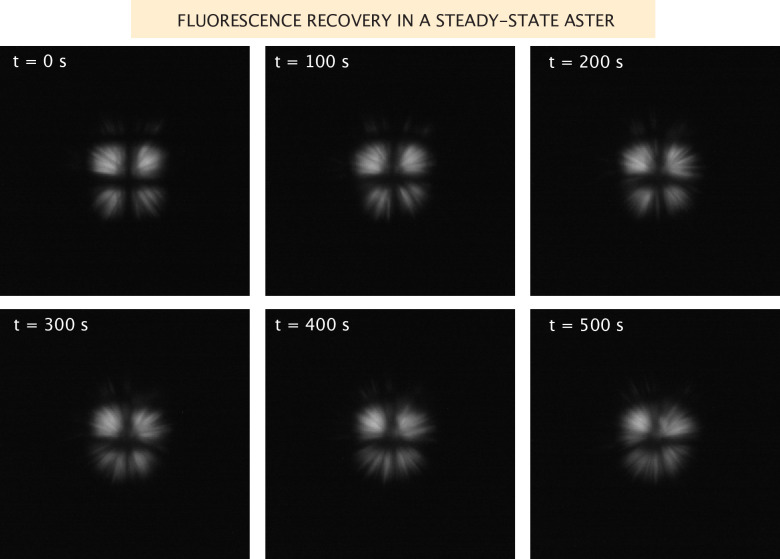
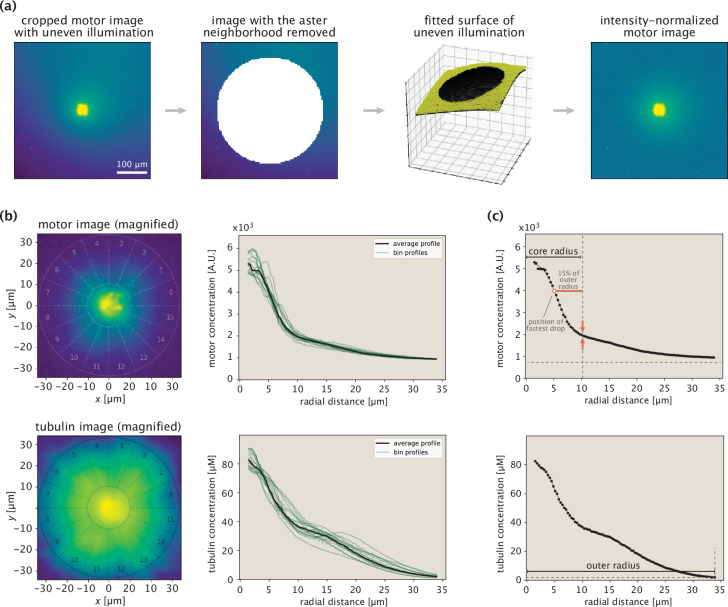
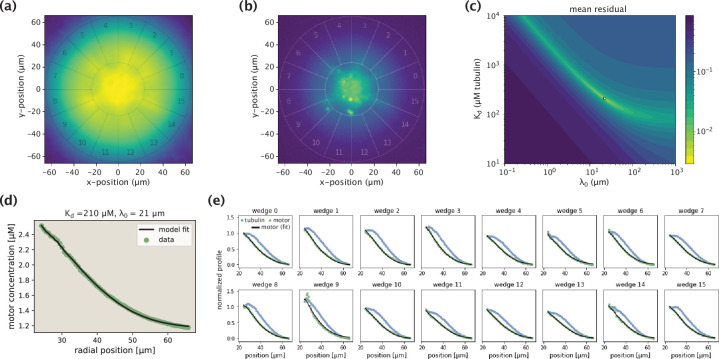
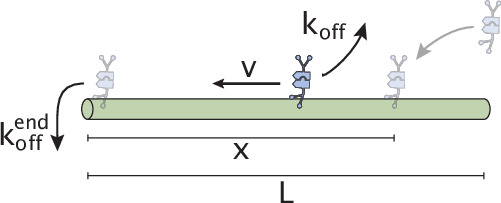
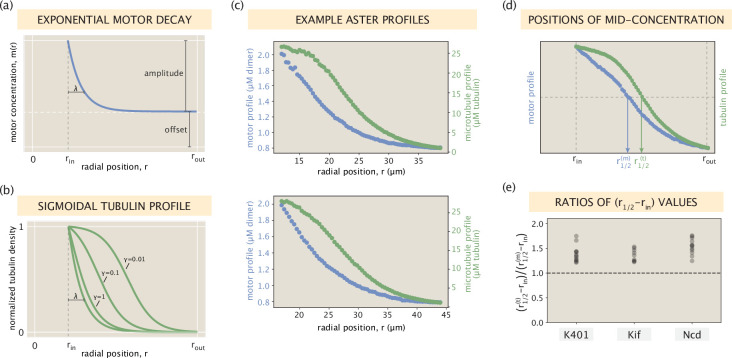
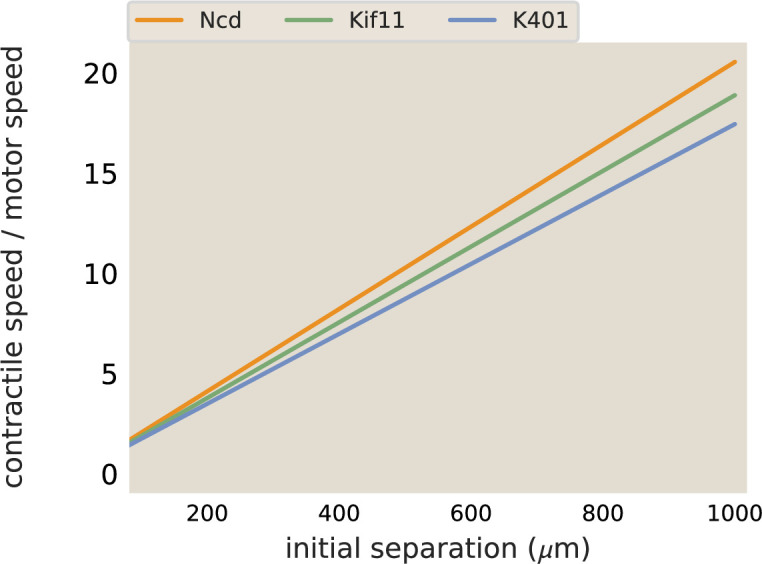
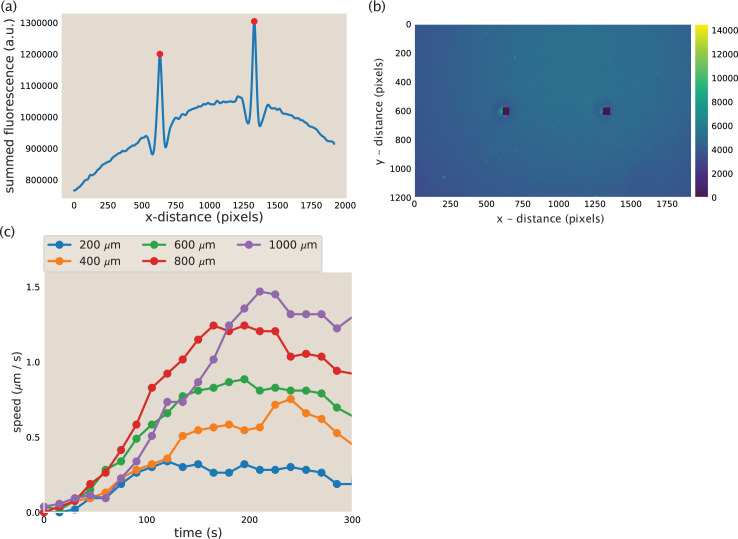
Active matter systems can generate highly ordered structures, avoiding equilibrium through the consumption of energy by individual constituents. How the microscopic parameters that characterize the active agents are translated to the observed mesoscopic properties of the assembly has remained an open question. These active systems are prevalent in living matter; for example, in cells, the cytoskeleton is organized into structures such as the mitotic spindle through the coordinated activity of many motor proteins walking along microtubules. Here, we investigate how the microscopic motor-microtubule interactions affect the coherent structures formed in a reconstituted motor-microtubule system. This question is of deeper evolutionary significance as we suspect motor and microtubule type contribute to the shape and size of resulting structures. We explore key parameters experimentally and theoretically, using a variety of motors with different speeds, processivities, and directionalities. We demonstrate that aster size depends on the motor used to create the aster, and develop a model for the distribution of motors and microtubules in steady-state asters that depends on parameters related to motor speed and processivity. Further, we show that network contraction rates scale linearly with the single-motor speed in quasi-one-dimensional contraction experiments. In all, this theoretical and experimental work helps elucidate how microscopic motor properties are translated to the much larger scale of collective motor-microtubule assemblies.
Keywords: active matter; aster; kinesin; microtubules; none; physics of living systems.
© 2023, Banks et al.
Conflict of interest statement
RB, VG, HL, SH, AI, TR, ZB, MT, RP No competing interests declared
Figures

Update of
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
