

Chemical Features of Polyanions Modulate Tau Aggregation and Conformational States
- PMID: 36753572
- PMCID: PMC9951223
- DOI: 10.1021/jacs.2c08004
Chemical Features of Polyanions Modulate Tau Aggregation and Conformational States
Abstract
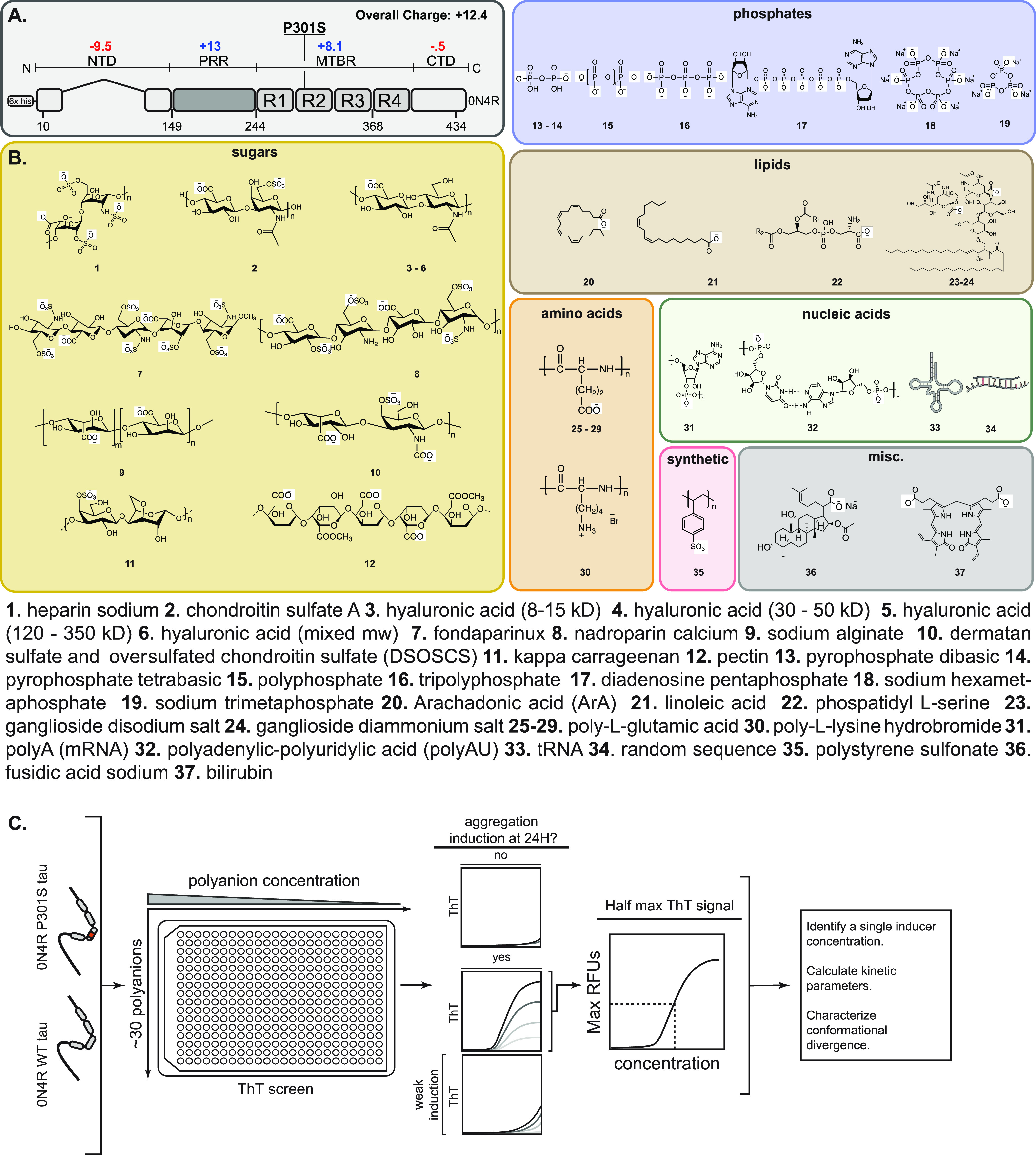
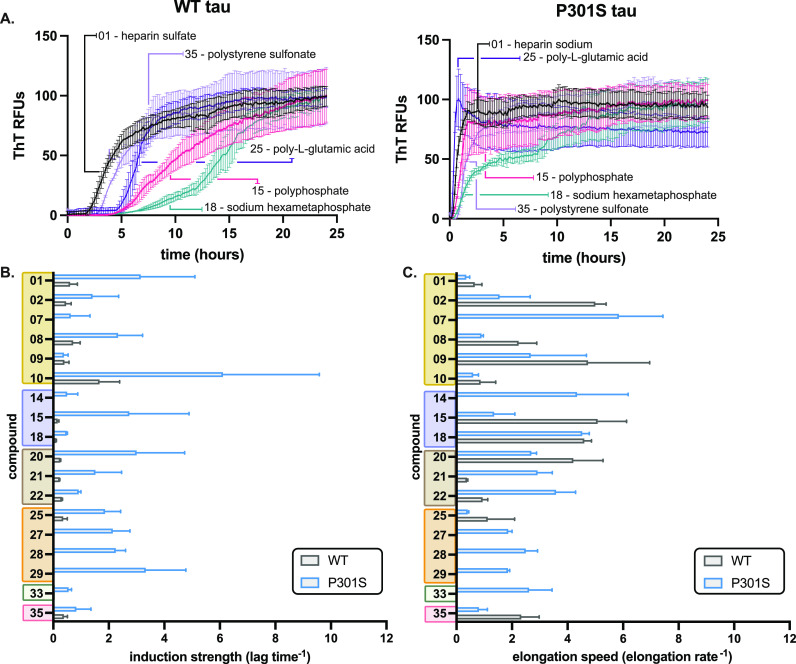
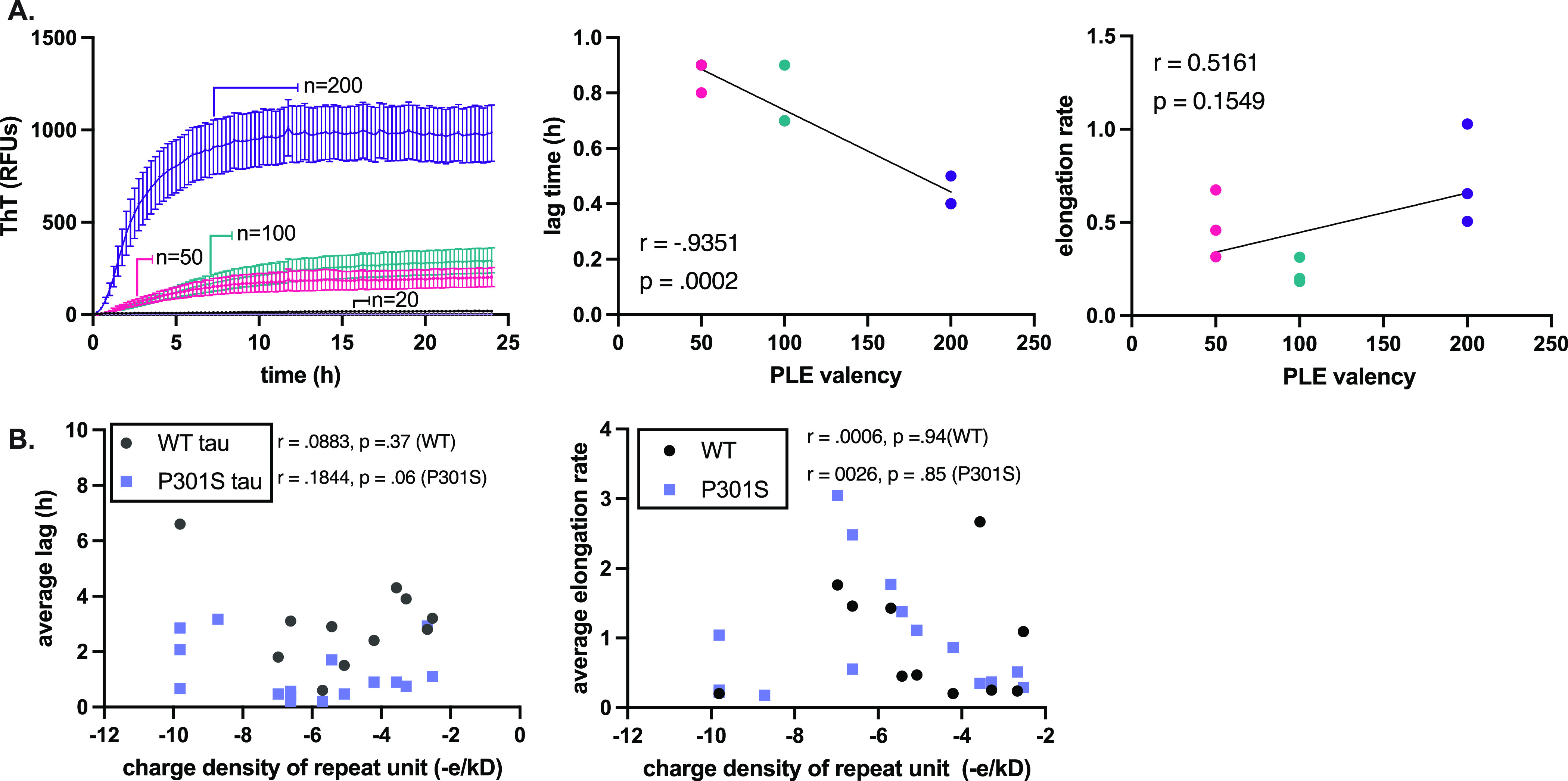
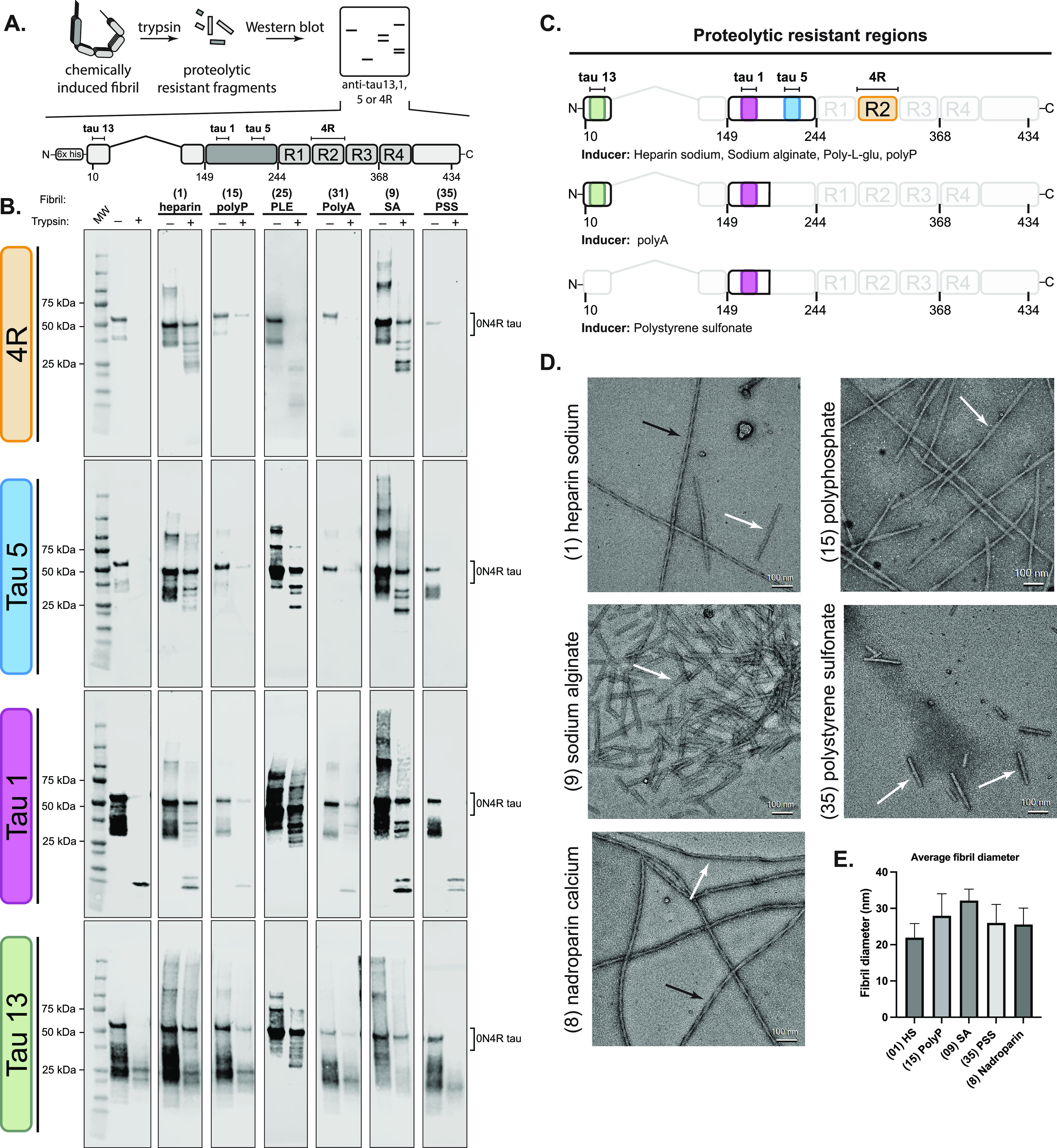
The aggregation of tau into insoluble fibrils is a defining feature of neurodegenerative tauopathies. However, tau has a positive overall charge and is highly soluble; so, polyanions, such as heparin, are typically required to promote its aggregation in vitro. There are dozens of polyanions in living systems, and it is not clear which ones might promote this process. Here, we systematically measure the ability of 37 diverse, anionic biomolecules to initiate tau aggregation using either wild-type (WT) tau or the disease-associated P301S mutant. We find that polyanions from many different structural classes can promote fibril formation and that P301S tau is sensitive to a greater number of polyanions (28/37) than WT tau (21/37). We also find that some polyanions preferentially reduce the lag time of the aggregation reactions, while others enhance the elongation rate, suggesting that they act on partially distinct steps. From the resulting structure-activity relationships, the valency of the polyanion seems to be an important chemical feature such that anions with low valency tend to be weaker aggregation inducers, even at the same overall charge. Finally, the identity of the polyanion influences fibril morphology based on electron microscopy and limited proteolysis. These results provide insights into the crucial role of polyanion-tau interactions in modulating tau conformational dynamics with implications for understanding the tau aggregation landscape in a complex cellular environment.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
-
- Holtzman D. M.; Carrillo M. C.; Hendrix J. A.; Bain L. J.; Catafau A. M.; Gault L. M.; Goedert M.; Mandelkow E.; Mandelkow E.-M.; Miller D. S.; Ostrowitzki S.; Polydoro M.; Smith S.; Wittmann M.; Hutton M. Tau: From Research to Clinical Development. Alzheimer’s Dementia 2016, 12, 1033–1039. 10.1016/j.jalz.2016.03.018. - DOI - PubMed
