

TET2 guards against unchecked BATF3-induced CAR T cell expansion
- PMID: 36755094
- PMCID: PMC10511001
- DOI: 10.1038/s41586-022-05692-z
TET2 guards against unchecked BATF3-induced CAR T cell expansion
Abstract
Further advances in cell engineering are needed to increase the efficacy of chimeric antigen receptor (CAR) and other T cell-based therapies1-5. As T cell differentiation and functional states are associated with distinct epigenetic profiles6,7, we hypothesized that epigenetic programming may provide a means to improve CAR T cell performance. Targeting the gene that encodes the epigenetic regulator ten-eleven translocation 2 (TET2)8 presents an interesting opportunity as its loss may enhance T cell memory9,10, albeit not cause malignancy9,11,12. Here we show that disruption of TET2 enhances T cell-mediated tumour rejection in leukaemia and prostate cancer models. However, loss of TET2 also enables antigen-independent CAR T cell clonal expansions that may eventually result in prominent systemic tissue infiltration. These clonal proliferations require biallelic TET2 disruption and sustained expression of the AP-1 factor BATF3 to drive a MYC-dependent proliferative program. This proliferative state is associated with reduced effector function that differs from both canonical T cell memory13,14 and exhaustion15,16 states, and is prone to the acquisition of secondary somatic mutations, establishing TET2 as a guardian against BATF3-induced CAR T cell proliferation and ensuing genomic instability. Our findings illustrate the potential of epigenetic programming to enhance T cell immunity but highlight the risk of unleashing unchecked proliferative responses.
© 2023. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interests.
Figures
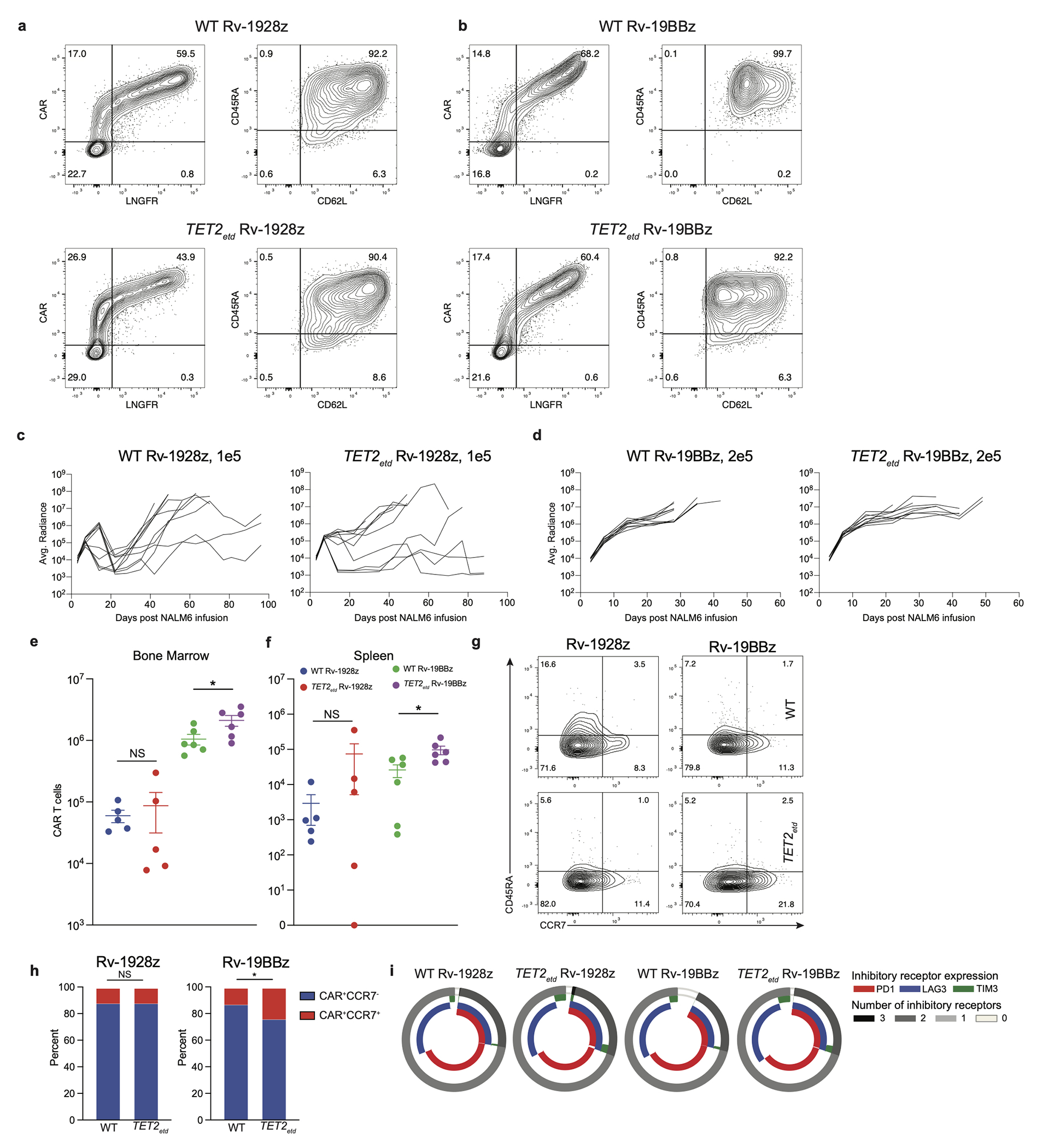
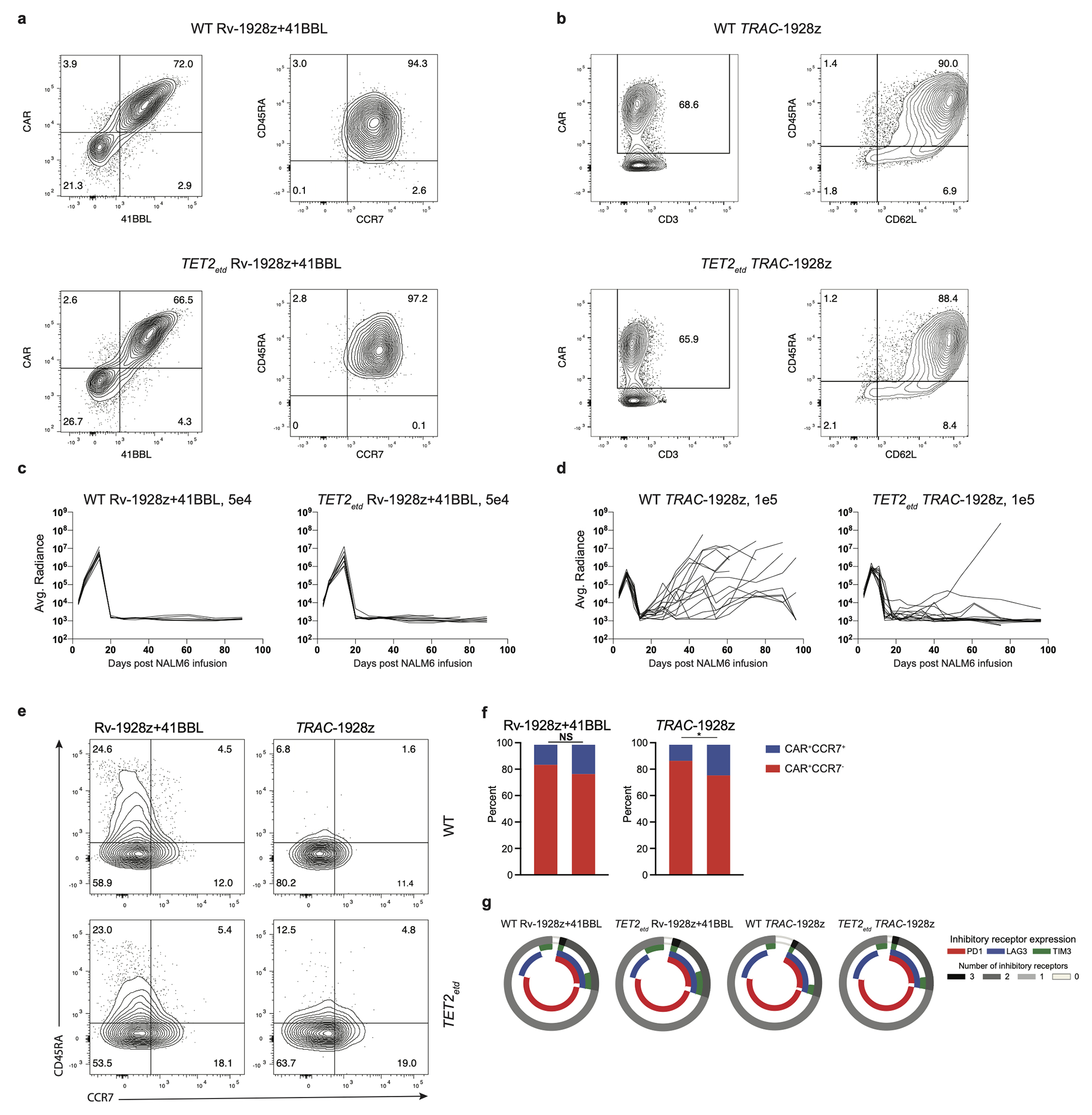
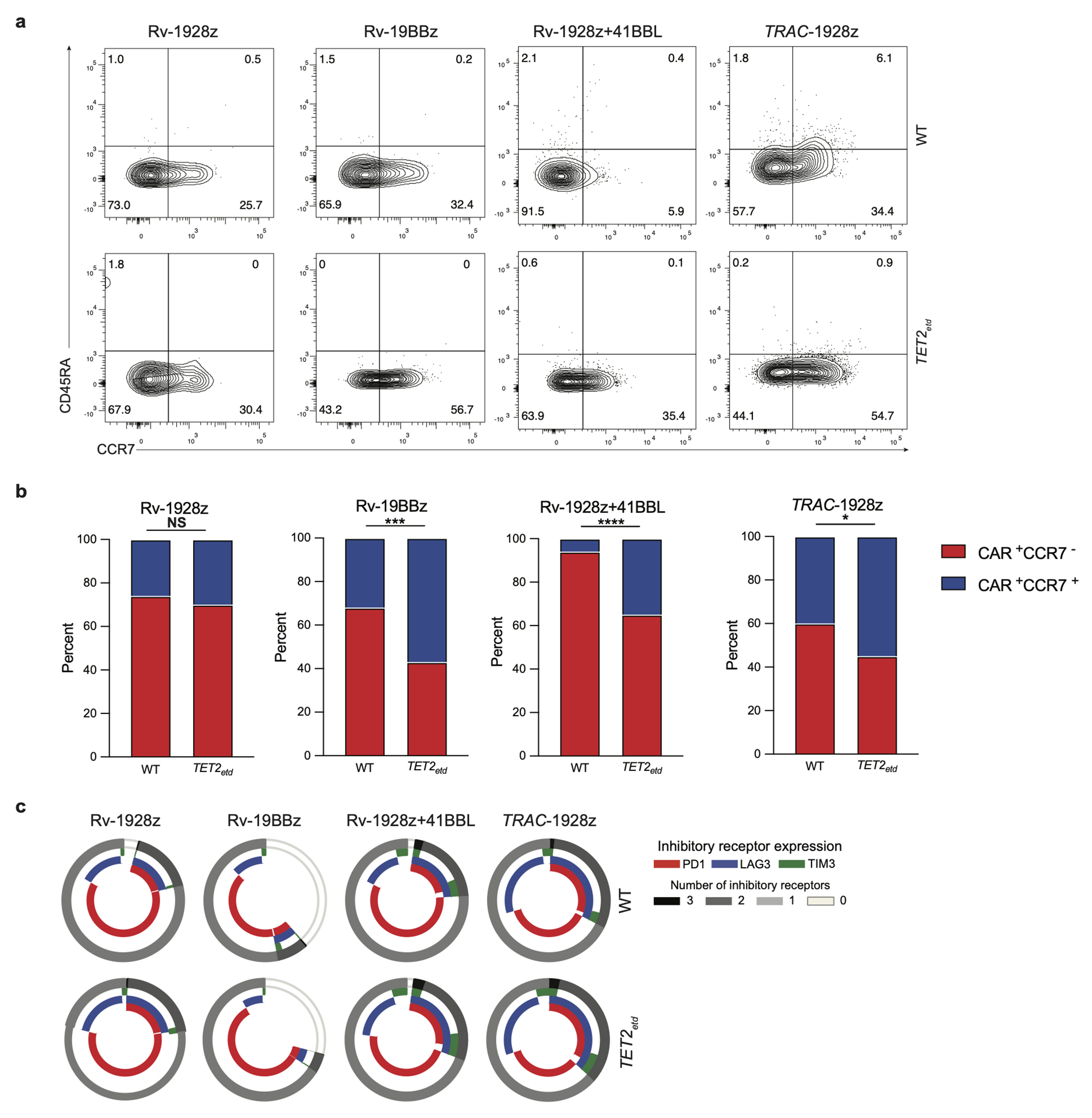
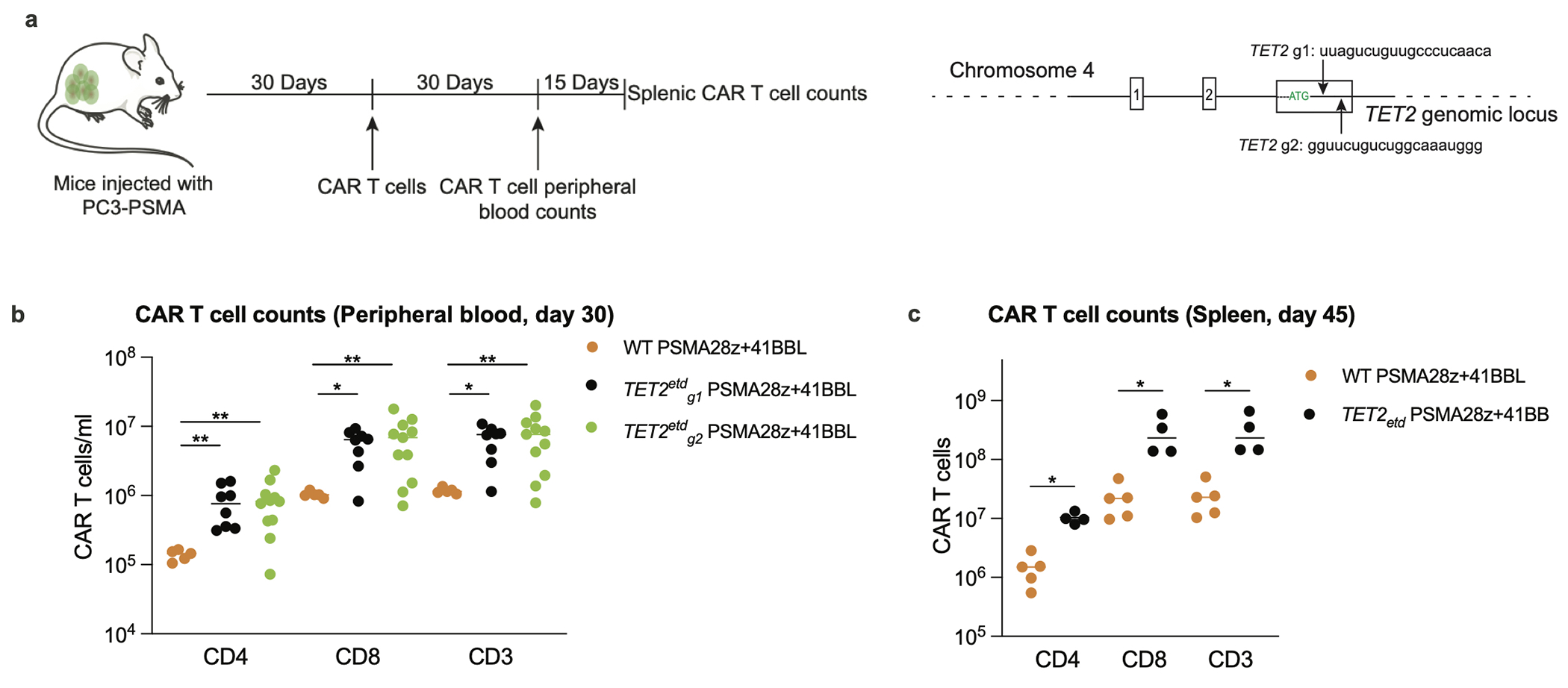
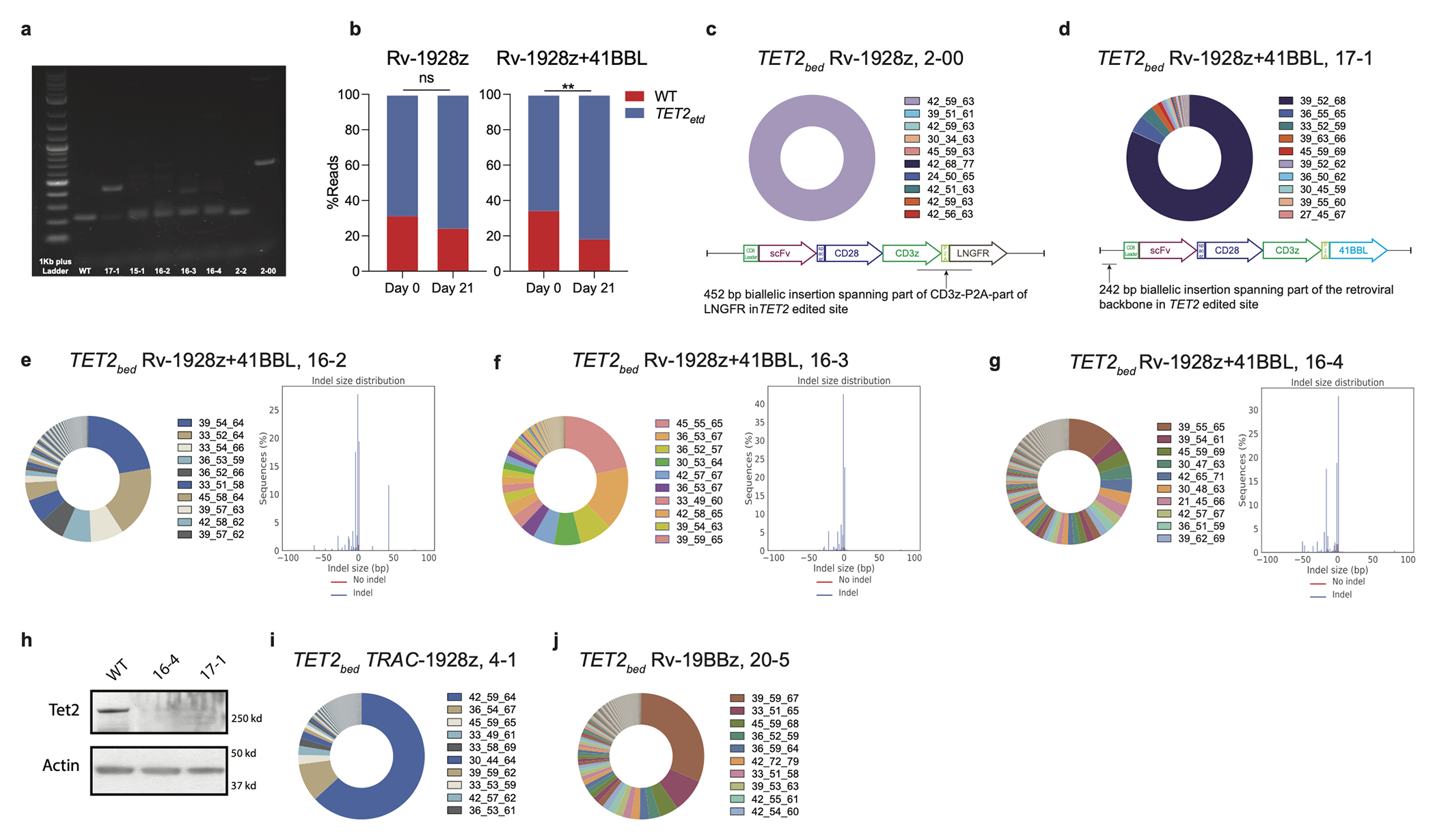
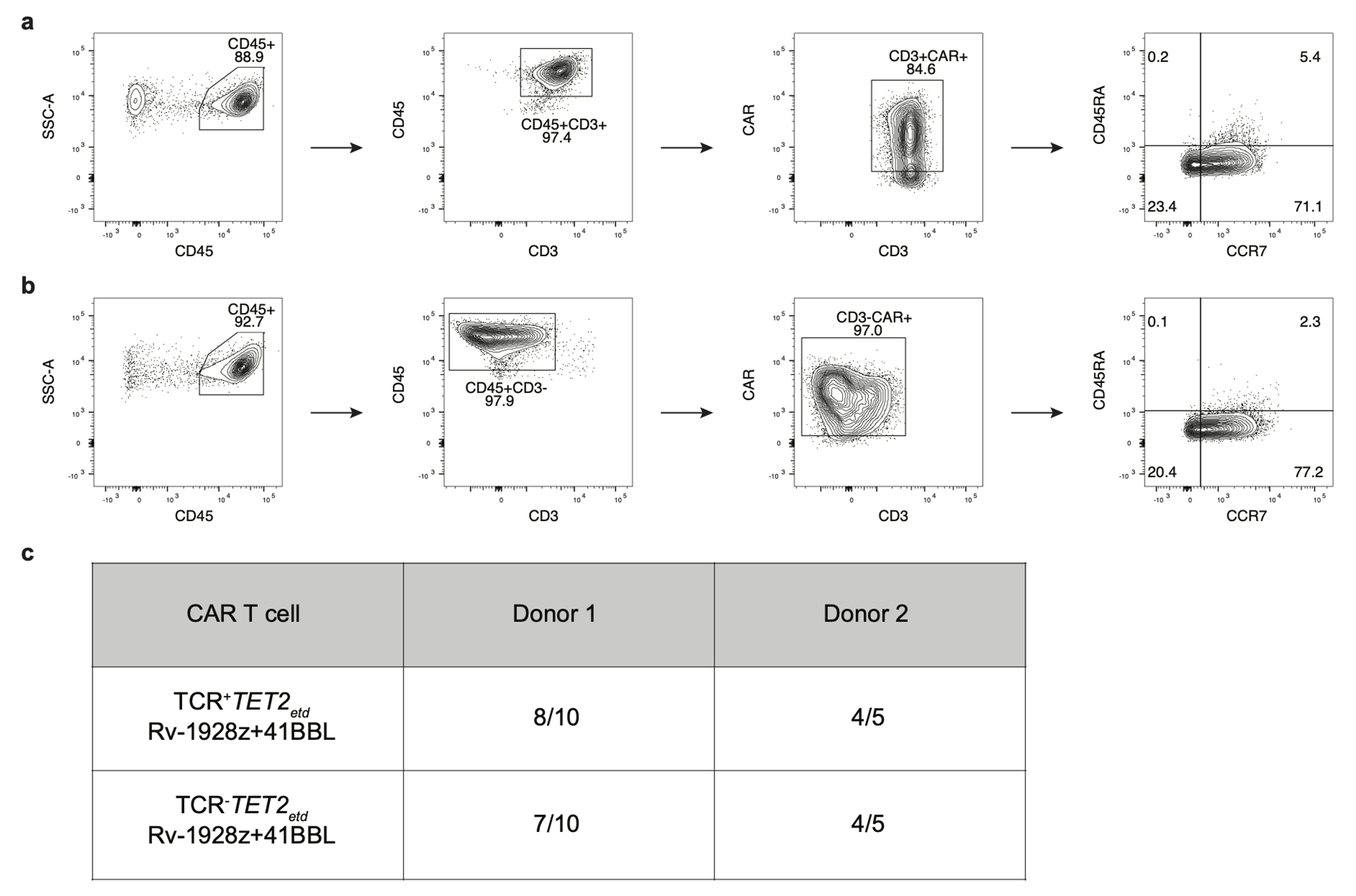
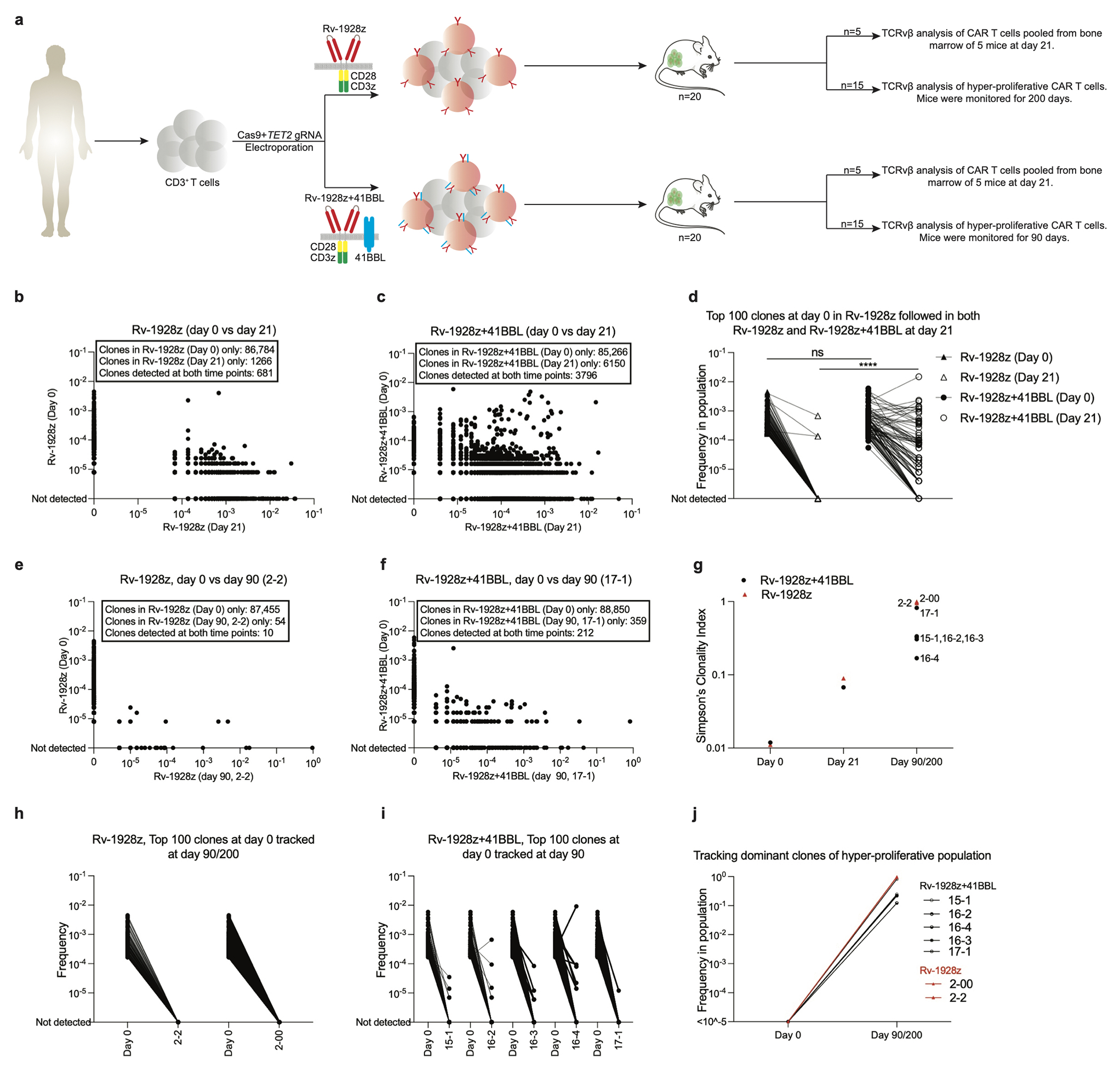
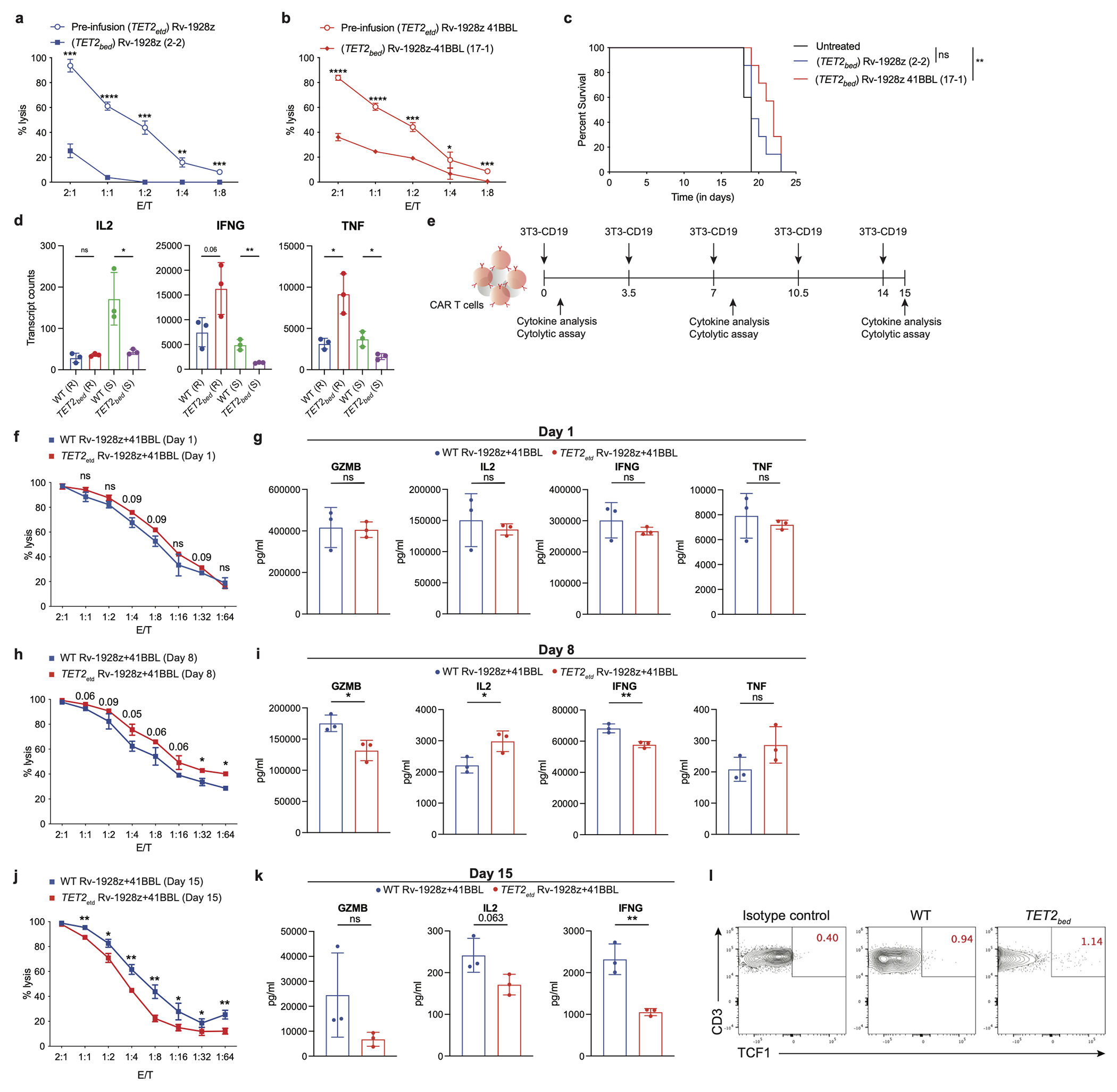
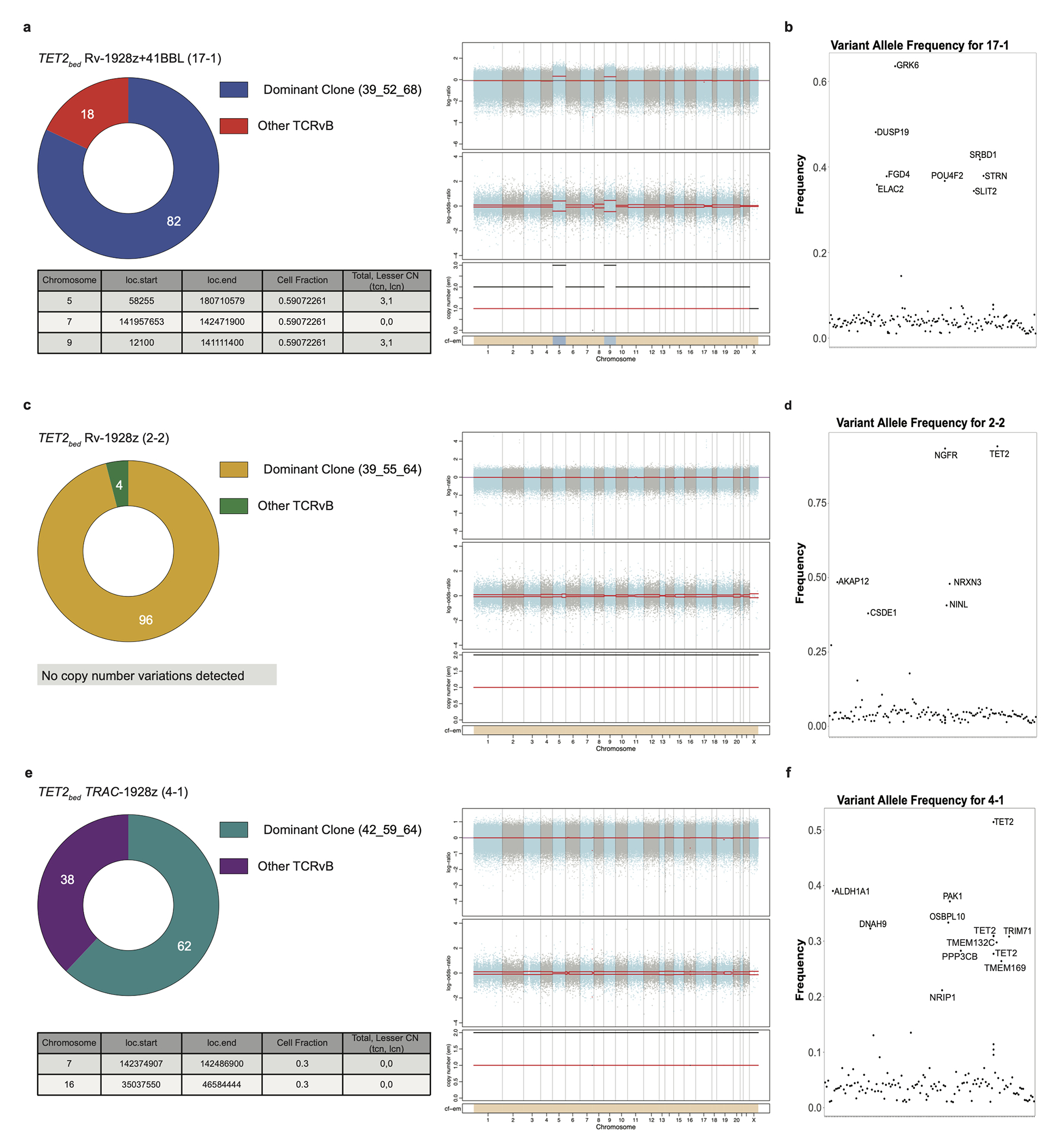
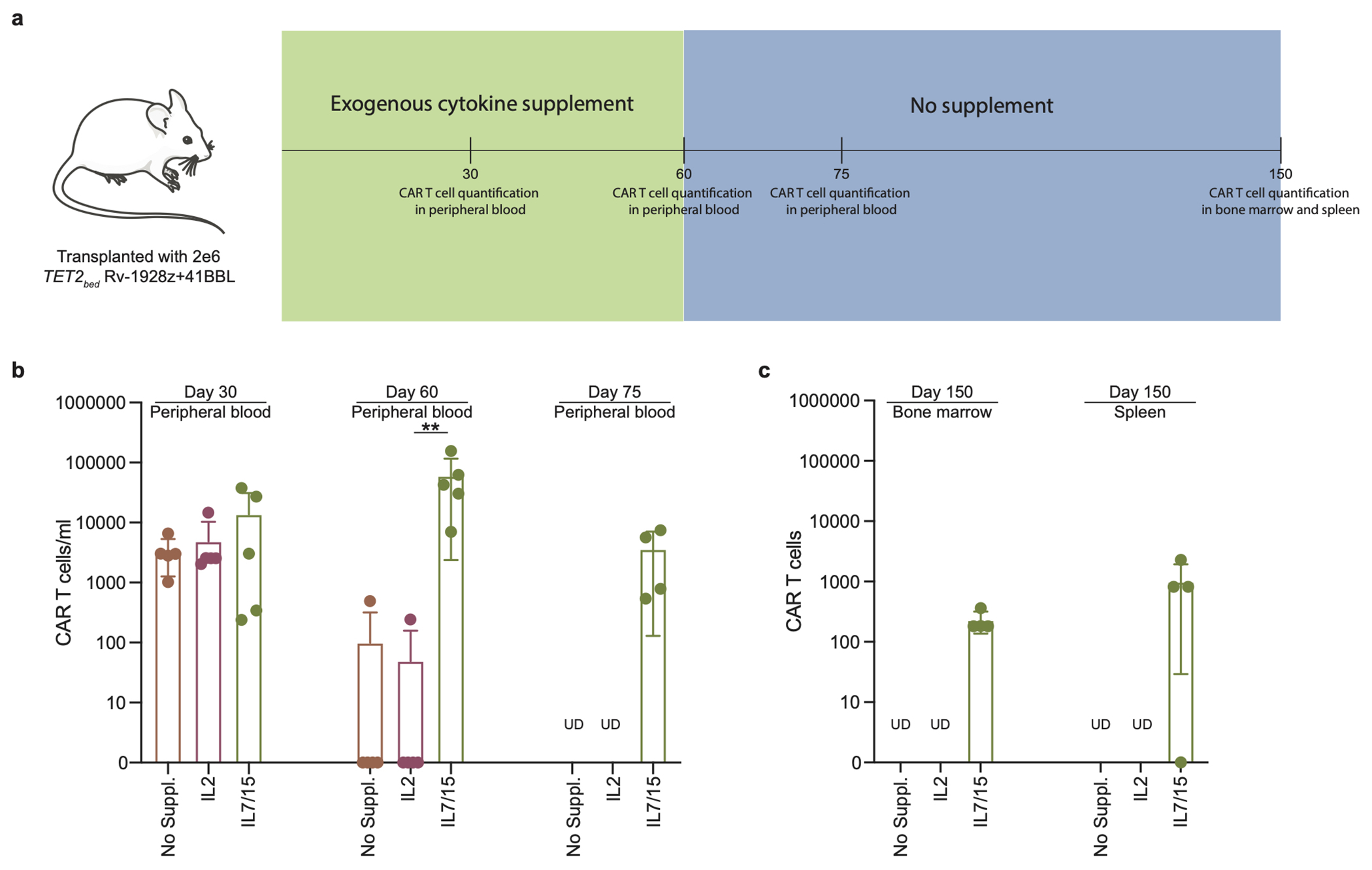
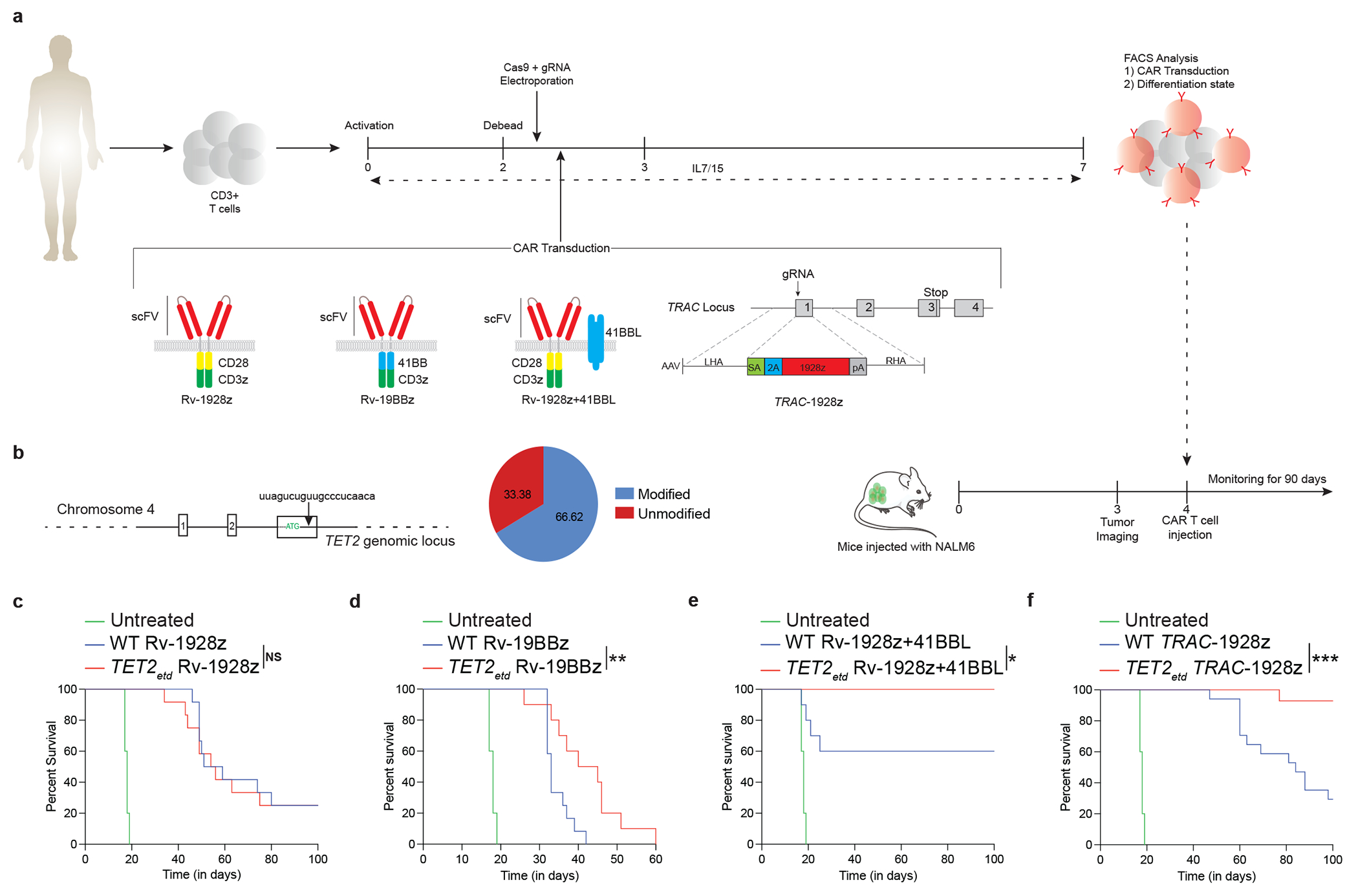
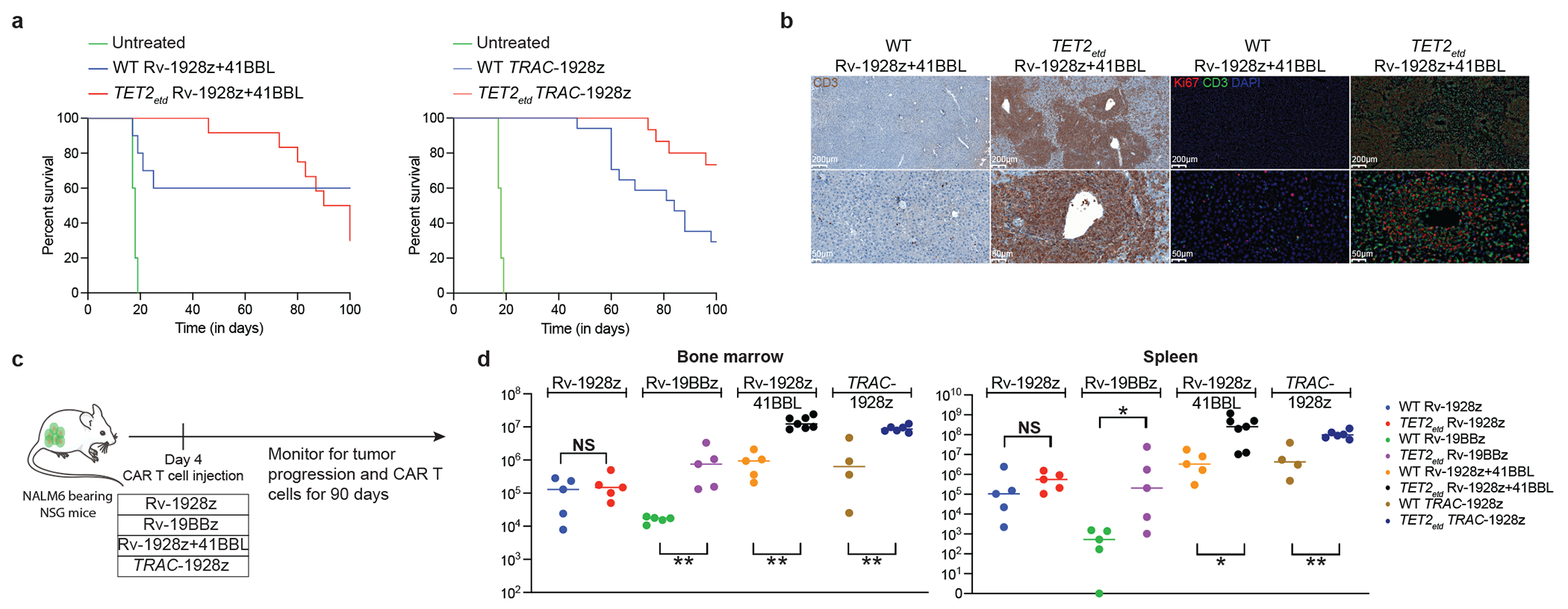
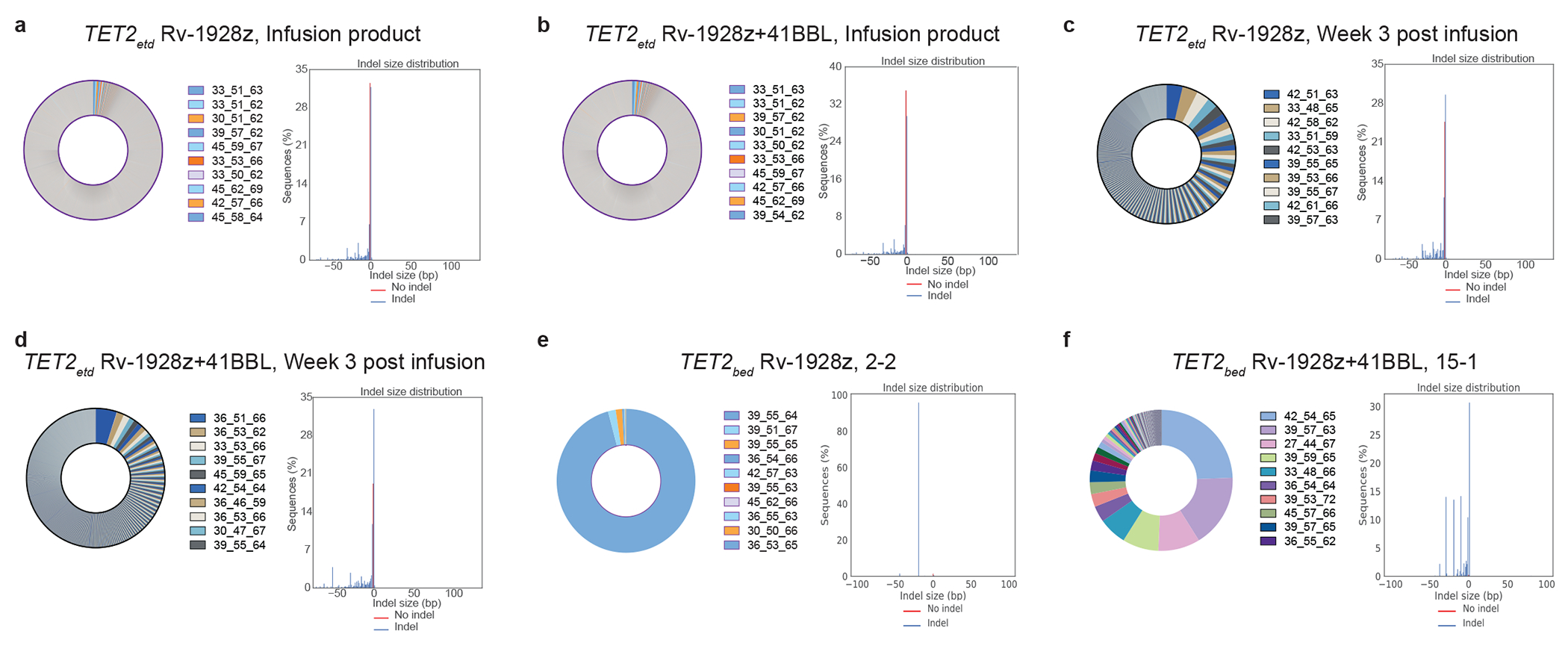
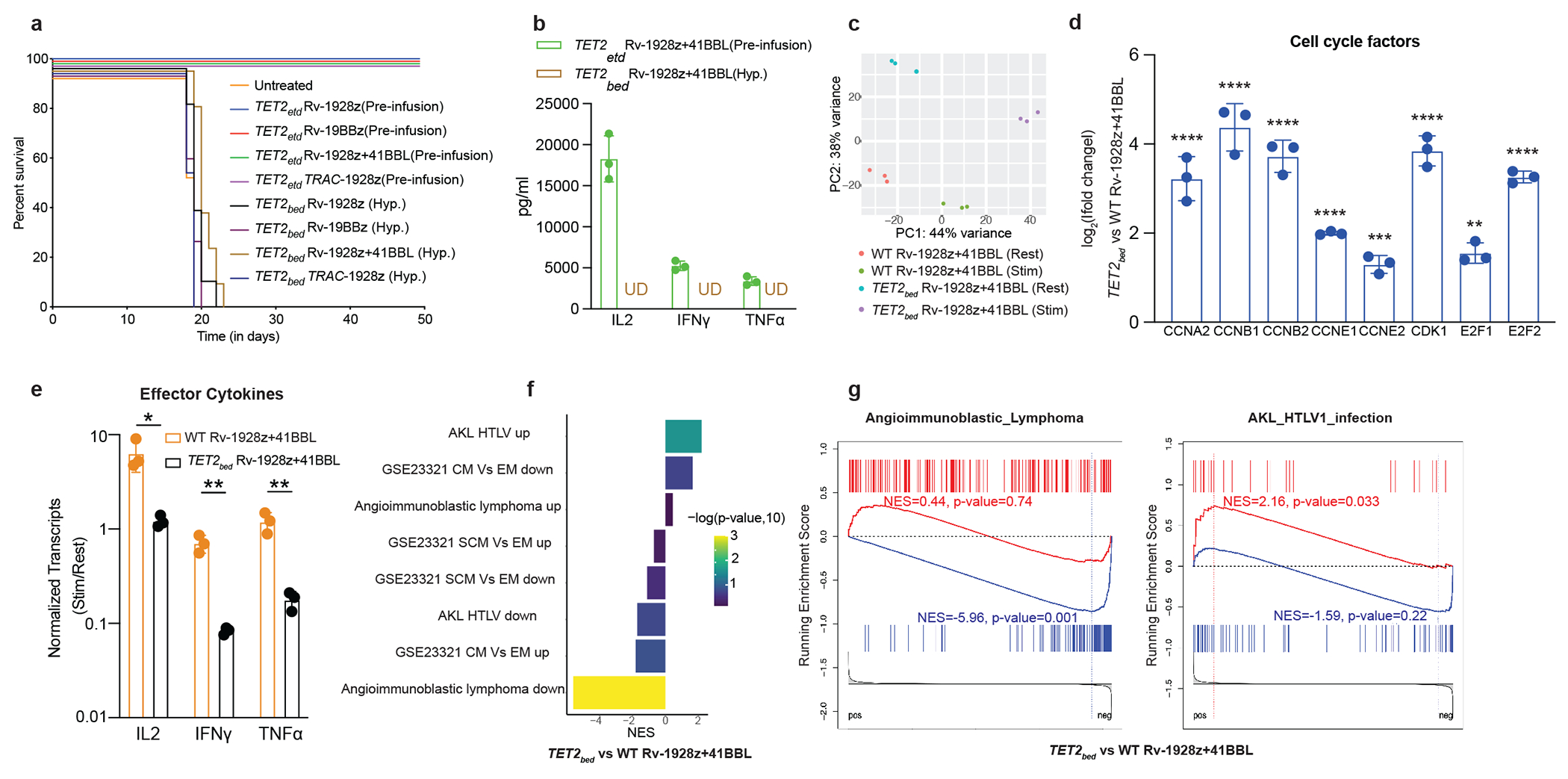
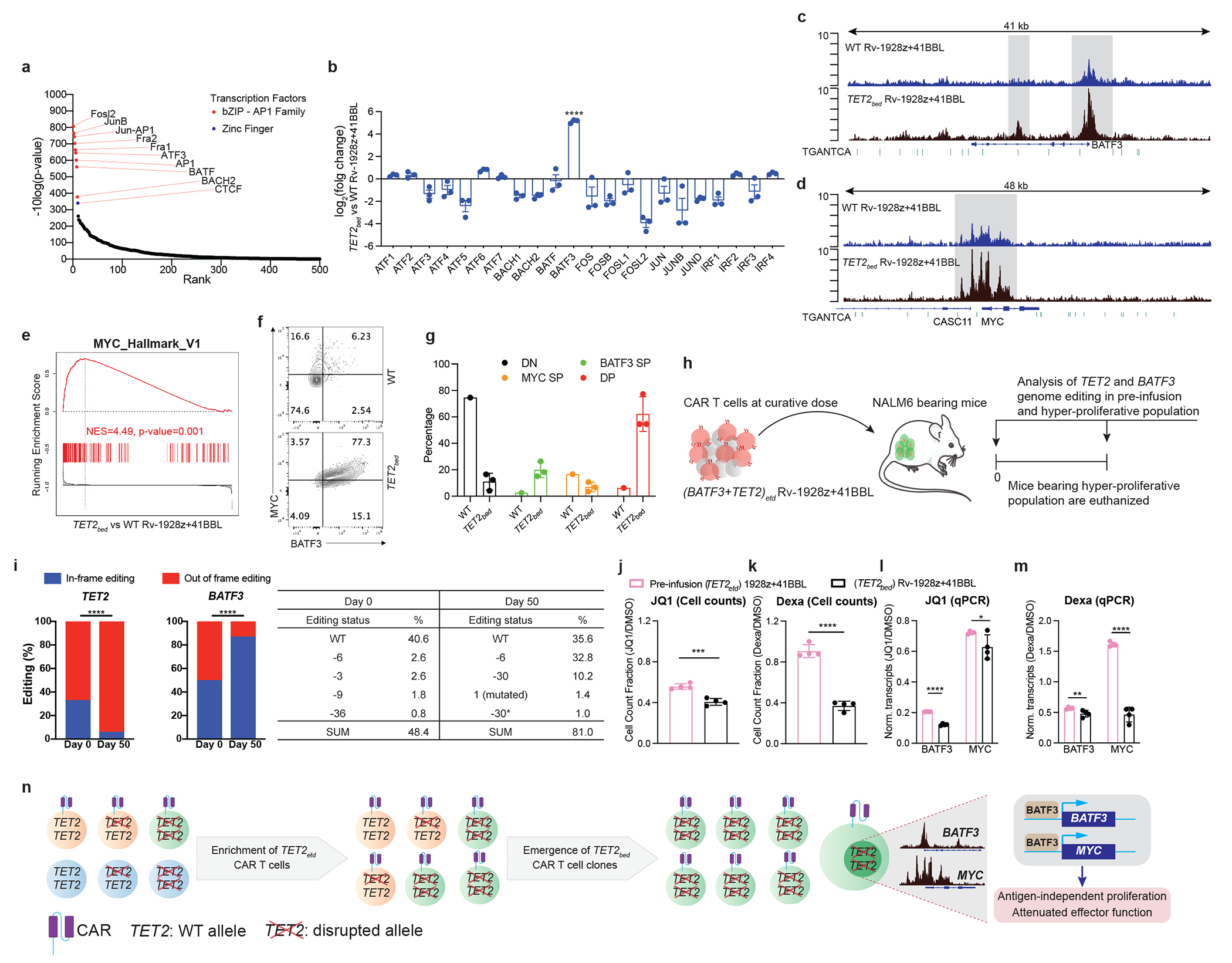
Comment in
-
TET2, tumor control, and CAR T cell hyperproliferation.Trends Cancer. 2023 Jul;9(7):521-523. doi: 10.1016/j.trecan.2023.04.009. Epub 2023 May 11. Trends Cancer. 2023. PMID: 37173187 Free PMC article.
References
Methods Reference
-
- Gallardo HF, Tan C, Ory D & Sadelain M Recombinant retroviruses pseudotyped with the vesicular stomatitis virus G glycoprotein mediate both stable gene transfer and pseudotransduction in human peripheral blood lymphocytes. Blood 90, 952–957 (1997). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
