

Human RSPO1 Mutation Represses Beige Adipocyte Thermogenesis and Contributes to Diet-Induced Adiposity
- PMID: 36755192
- PMCID: PMC10131814
- DOI: 10.1002/advs.202207152
Human RSPO1 Mutation Represses Beige Adipocyte Thermogenesis and Contributes to Diet-Induced Adiposity
Abstract
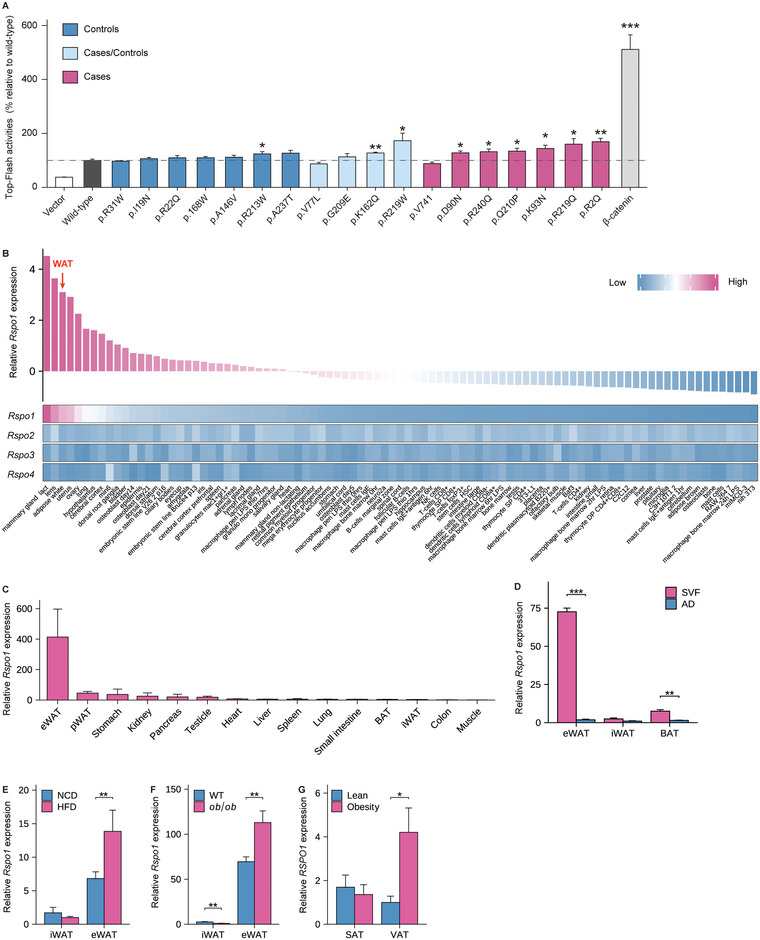
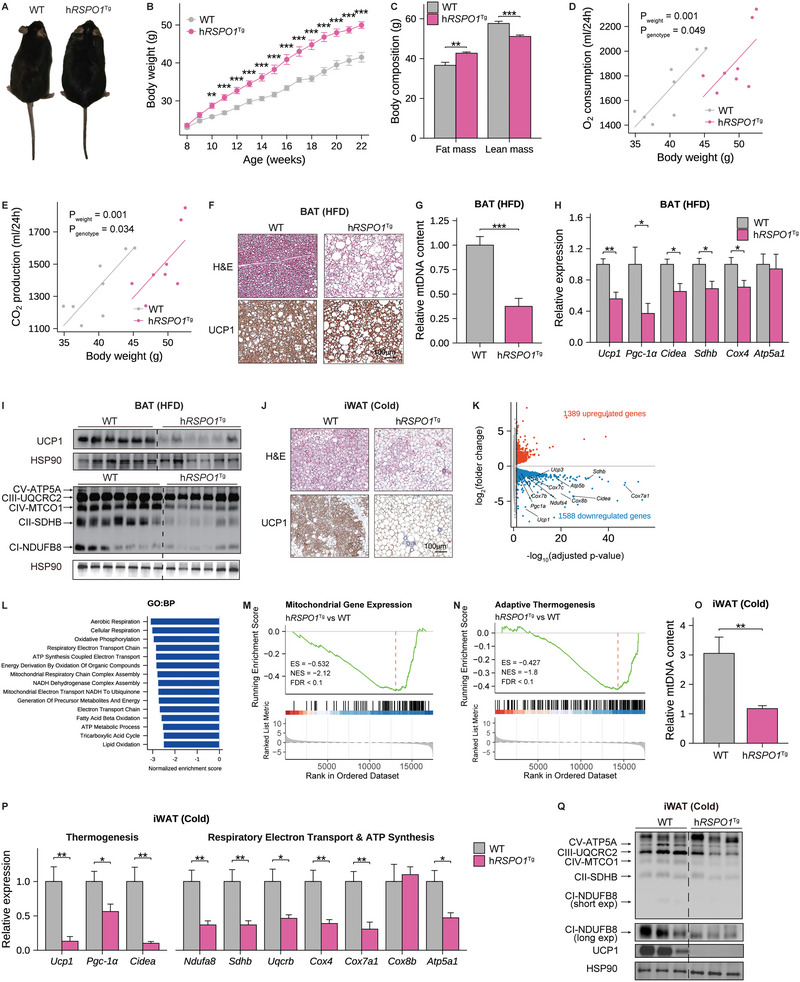
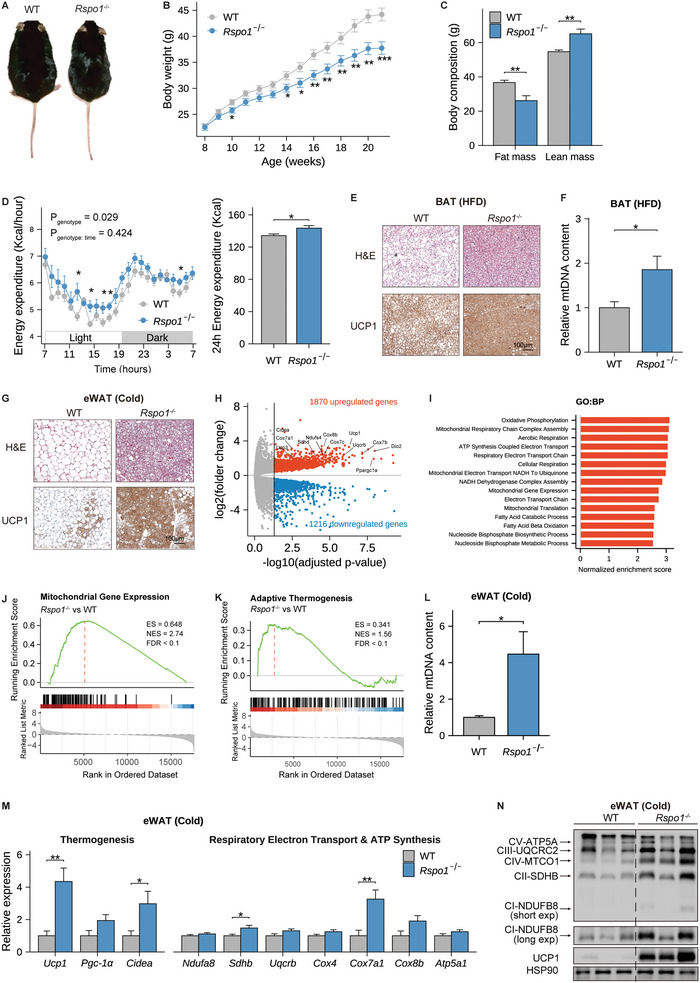
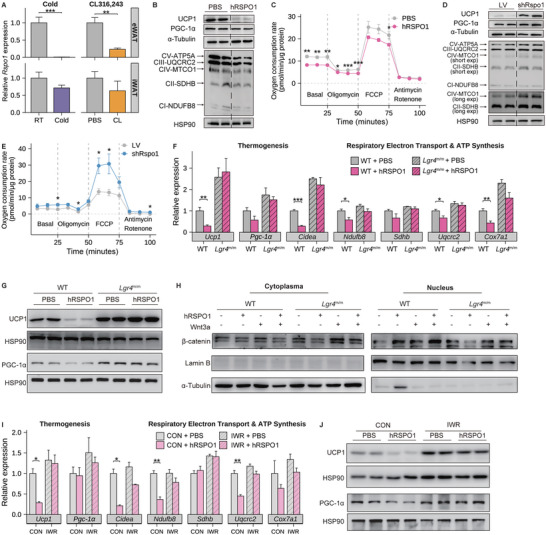
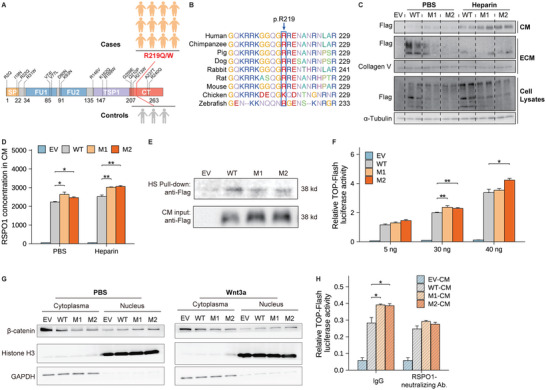
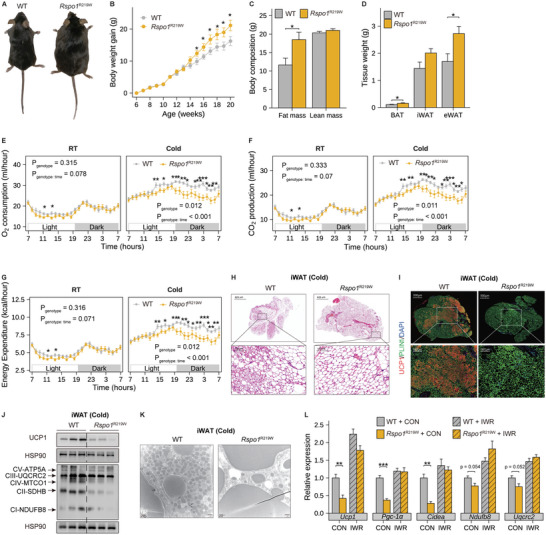
Recent genetic evidence has linked WNT downstream mutations to fat distribution. However, the roles of WNTs in human obesity remain unclear. Here, the authors screen all Wnt-related paracrine factors in 1994 obese cases and 2161 controls using whole-exome sequencing (WES) and identify that 12 obese patients harbor the same mutations in RSPO1 (p.R219W/Q) predisposing to human obesity. RSPO1 is predominantly expressed in visceral fat, primarily in the fibroblast cluster, and is increased with adiposity. Mice overexpressing human RSPO1 in adipose tissues develop obesity under a high-fat diet (HFD) due to reduced brown/beige fat thermogenesis. In contrast, Rspo1 ablation resists HFD-induced adiposity by increasing thermogenesis. Mechanistically, RSPO1 overexpression or administration significantly inhibits adipocyte mitochondrial respiration and thermogenesis via LGR4-Wnt/β-catenin signaling pathway. Importantly, humanized knockin mice carrying the hotspot mutation (p.R219W) display suppressed thermogenesis and recapitulate the adiposity feature of obese carriers. The mutation disrupts RSPO1's electrostatic interaction with the extracellular matrix, leading to excessive RSPO1 release that activates LGR4-Wnt/β-catenin signaling and attenuates thermogenic capacity in differentiated beige adipocytes. Therefore, these findings identify that gain-of-function mutations and excessive expression of RSPO1, acting as a paracrine Wnt activator, suppress fat thermogenesis and contribute to obesity in humans.
Keywords: RSPO1; Wnt signaling; obesity; pathogenic gene; thermogenesis; whole-exome sequencing.
© 2023 The Authors. Advanced Science published by Wiley-VCH GmbH.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- a) van Marken Lichtenbelt W. D., Vanhommerig J. W., Smulders N. M., Drossaerts J. M. A. F. L., Kemerink G. J., Bouvy N. D., Schrauwen P., Teule G. J. J., N. Engl. J. Med. 2009, 360, 1500; - PubMed
- b) Din M. U., Saari T., Raiko J., Kudomi N., Maurer S. F., Lahesmaa M., Fromme T., Amri E.‐Z., Klingenspor M., Solin O., Nuutila P., Virtanen K. A., Cell Metab. 2018, 28, 207. - PubMed
-
- Giordano A., Frontini A., Cinti S., Nat. Rev. Drug Discovery 2016, 15, 405. - PubMed
-
- a) Feldmann H. M., Golozoubova V., Cannon B., Nedergaard J., Cell Metab. 2009, 9, 203; - PubMed
- b) Mano‐Otagiri A., Iwasaki‐Sekino A., Nemoto T., Ohata H., Shuto Y., Nakabayashi H., Sugihara H., Oikawa S., Shibasaki T., Regul. Pept. 2010, 160, 81; - PubMed
- c) Berg F., Gustafson U., Andersson L., PLoS Genet. 2006, 2, e129; - PMC - PubMed
- d) Zheng Q., Lin J., Huang J., Zhang H., Zhang R., Zhang X., Cao C., Hambly C., Qin G., Yao J., Song R., Jia Q., Wang X., Li Y., Zhang N., Piao Z., Ye R., Speakman J. R., Wang H., Zhou Q., Wang Y., Jin W., Zhao J., Proc. Natl. Acad. Sci. USA 2017, 114, E9474. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- 2022YFC2505201/National Key Research and Development Program of China
- 2021YFA1301103/National Key Research and Development Program of China
- 91957124/National Natural Science Foundation of China
- 92157204/National Natural Science Foundation of China
- 82088102/National Natural Science Foundation of China
- 82250901/National Natural Science Foundation of China
- 82050007/National Natural Science Foundation of China
- 81930021/National Natural Science Foundation of China
- 81822009/National Natural Science Foundation of China
- 91857205/National Natural Science Foundation of China
- 81730023/National Natural Science Foundation of China
- 81870585/National Natural Science Foundation of China
- Innovative research team of high-level local universities in Shanghai
- 20161306/Shanghai Municipal Education Commission-Gaofeng Clinical Medicine
- 20171903/Shanghai Municipal Education Commission-Gaofeng Clinical Medicine
- 2018BR01/Outstanding Academic Leader Program of Shanghai Municipal Health Commission
- 21JC1404400/Science and Technology Commission of Shanghai Municipality
- 20XD1403200/Program of Shanghai Academic/Technology Research Leader
LinkOut - more resources
Full Text Sources
Molecular Biology Databases