

Melanophages give rise to hyperreflective foci in AMD, a disease-progression marker
- PMID: 36755326
- PMCID: PMC9906876
- DOI: 10.1186/s12974-023-02699-9
Melanophages give rise to hyperreflective foci in AMD, a disease-progression marker
Abstract
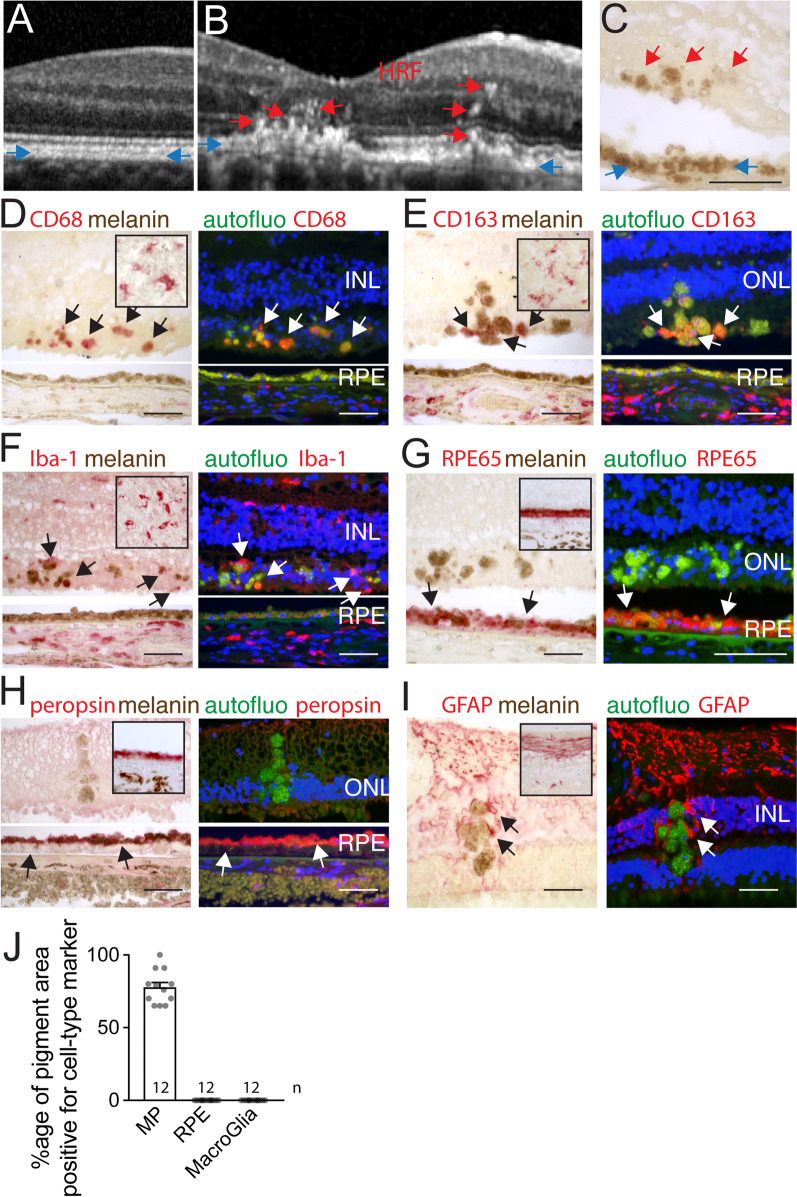
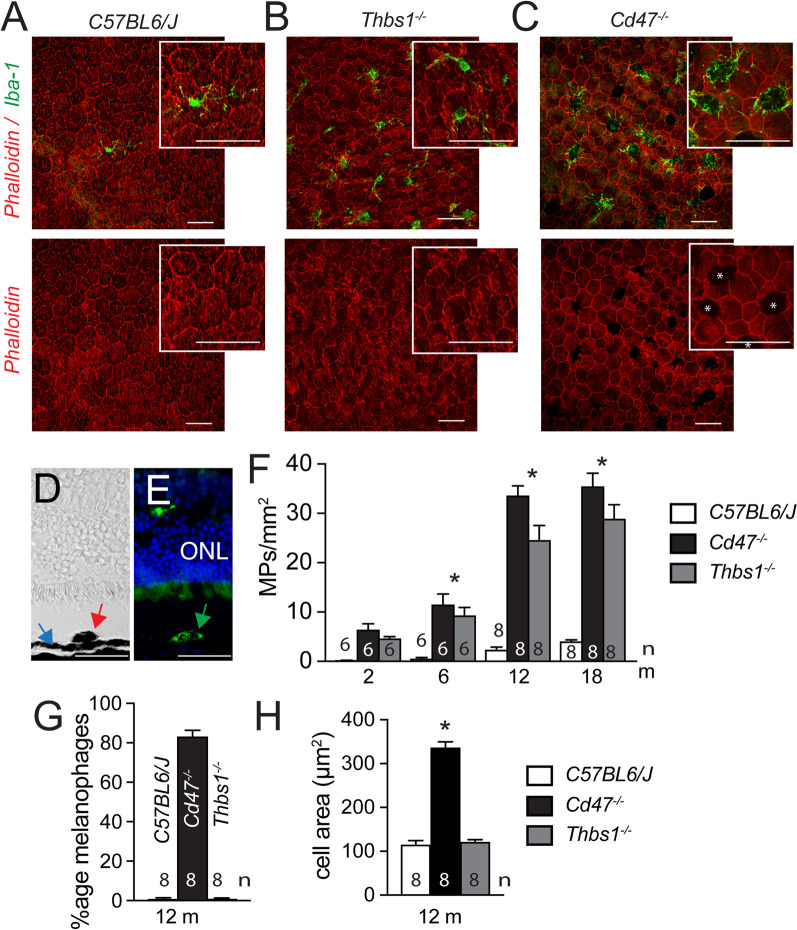
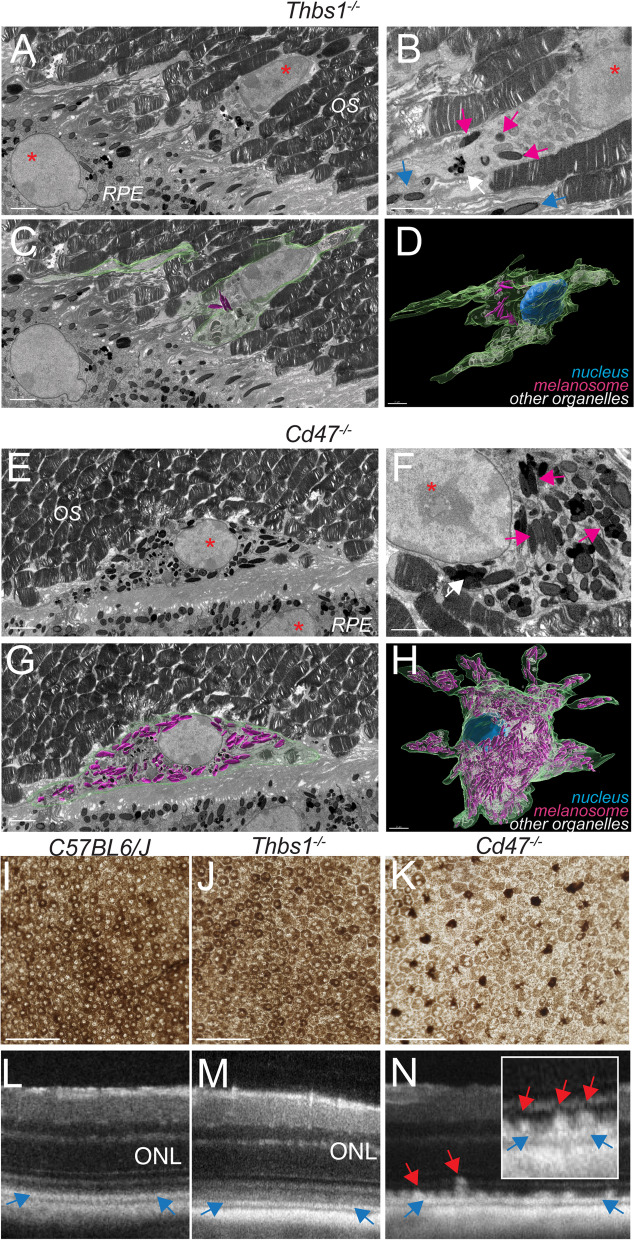
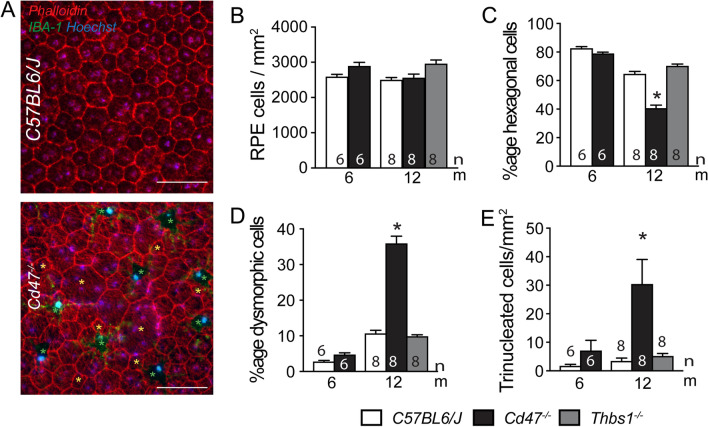
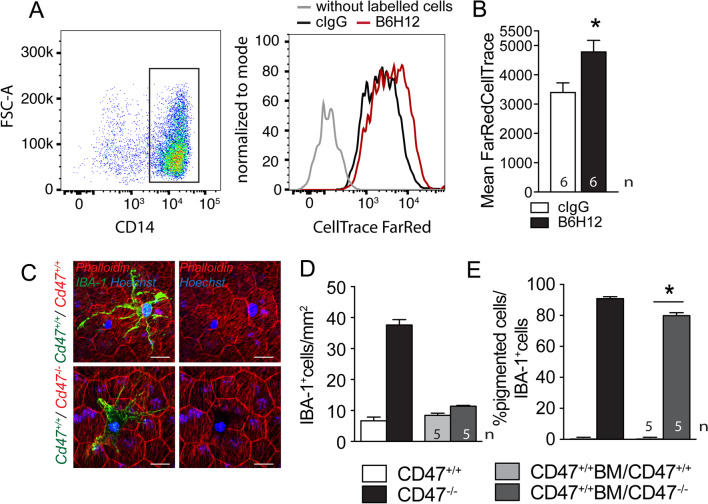
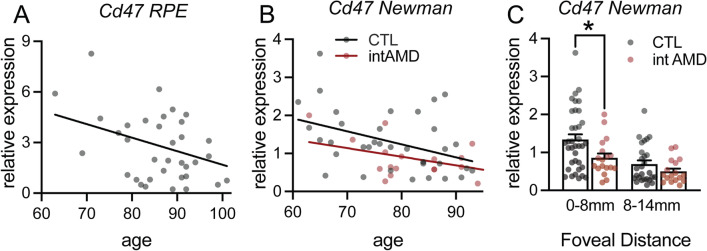
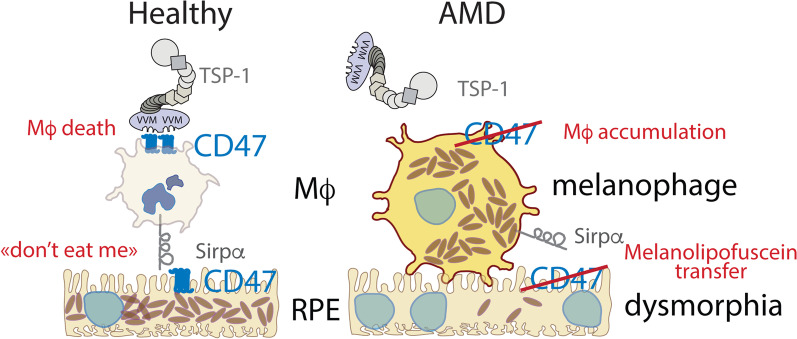
Retinal melanosome/melanolipofuscin-containing cells (MCCs), clinically visible as hyperreflective foci (HRF) and a highly predictive imaging biomarker for the progression of age-related macular degeneration (AMD), are widely believed to be migrating retinal pigment epithelial (RPE) cells. Using human donor tissue, we identify the vast majority of MCCs as melanophages, melanosome/melanolipofuscin-laden mononuclear phagocytes (MPs). Using serial block-face scanning electron microscopy, RPE flatmounts, bone marrow transplantation and in vitro experiments, we show how retinal melanophages form by the transfer of melanosomes from the RPE to subretinal MPs when the "don't eat me" signal CD47 is blocked. These melanophages give rise to hyperreflective foci in Cd47-/--mice in vivo, and are associated with RPE dysmorphia similar to intermediate AMD. Finally, we show that Cd47 expression in human RPE declines with age and in AMD, which likely participates in melanophage formation and RPE decline. Boosting CD47 expression in AMD might protect RPE cells and delay AMD progression.
Keywords: Age-related macular degeneration; CD47; Macrophage; Neuroinflammation.
© 2023. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Wong WL, Su X, Li X, Cheung CM, Klein R, Cheng CY, Wong TY. Global prevalence of age-related macular degeneration and disease burden projection for 2020 and 2040: a systematic review and meta-analysis. Lancet Glob Health. 2014;2:e106–116. - PubMed
-
- Fleckenstein M, Keenan TDL, Guymer RH, Chakravarthy U, Schmitz-Valckenberg S, Klaver CC, Wong WT, Chew EY. Age-related macular degeneration. Nat Rev Dis Primers. 2021;7:31. - PubMed
-
- Guillonneau X, Eandi CM, Paques M, Sahel JA, Sapieha P, Sennlaub F. On phagocytes and macular degeneration. Prog Retin Eye Res. 2017;61:98–128. - PubMed
-
- Klein R, Peto T, Bird A, Vannewkirk MR. The epidemiology of age-related macular degeneration. Am J Ophthalmol. 2004;137:486–495. - PubMed
-
- Wang JJ, Foran S, Smith W, Mitchell P. Risk of age-related macular degeneration in eyes with macular drusen or hyperpigmentation: the Blue Mountains Eye Study cohort. Arch Ophthalmol. 2003;121:658–663. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
