

A terpene nucleoside from M. tuberculosis induces lysosomal lipid storage in foamy macrophages
- PMID: 36757797
- PMCID: PMC10014106
- DOI: 10.1172/JCI161944
A terpene nucleoside from M. tuberculosis induces lysosomal lipid storage in foamy macrophages
Abstract
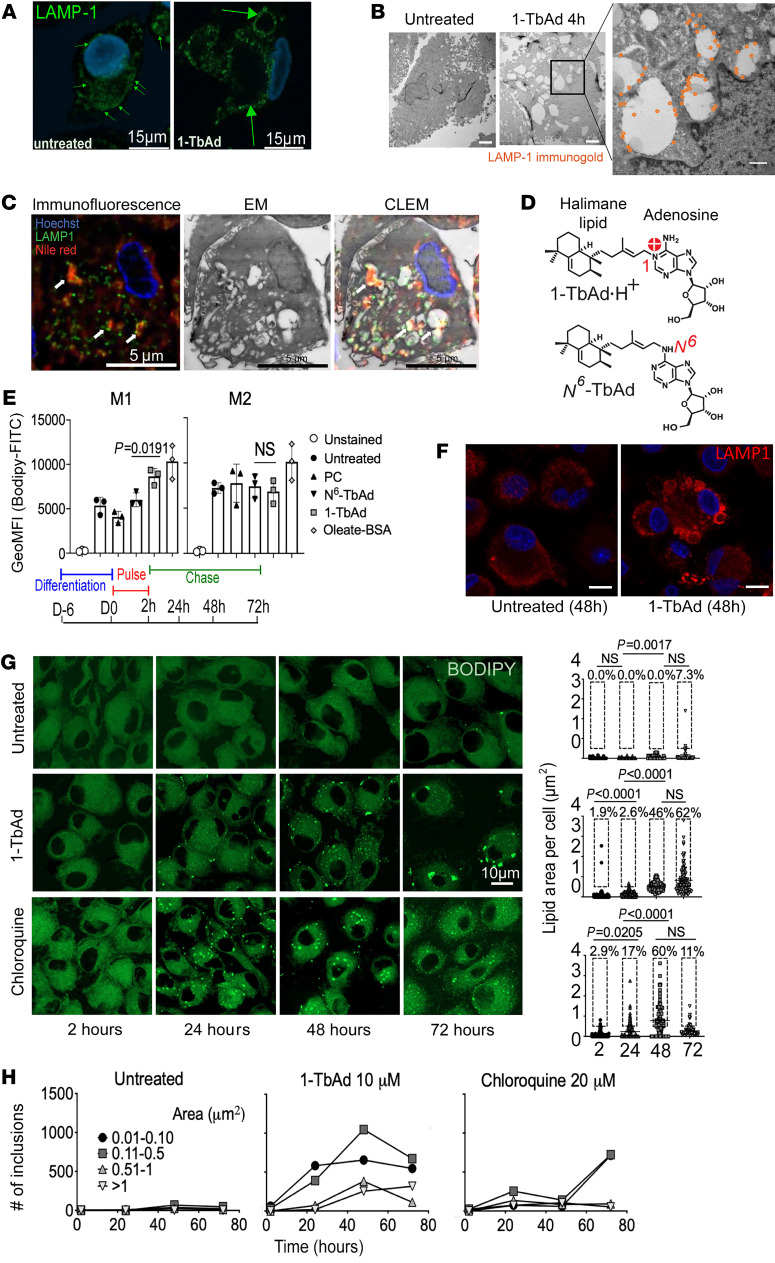
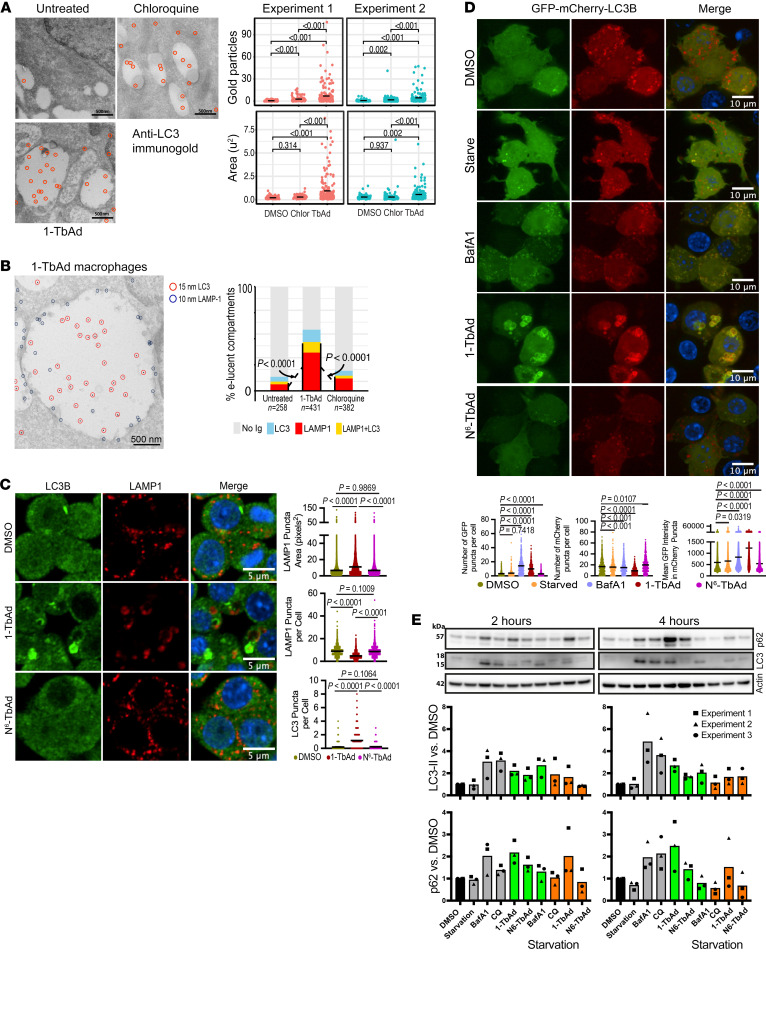
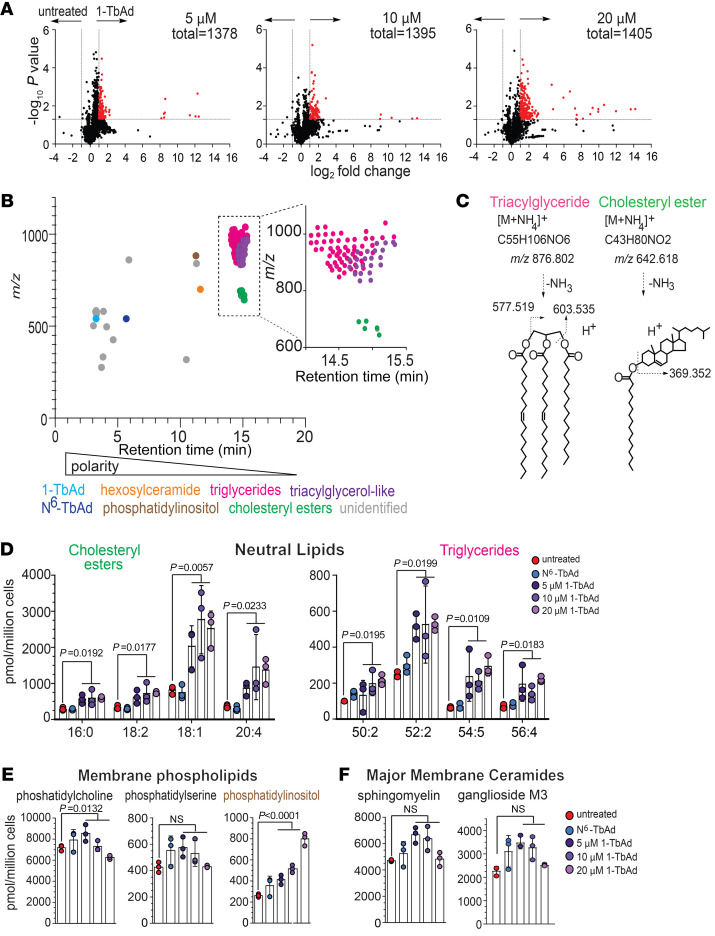
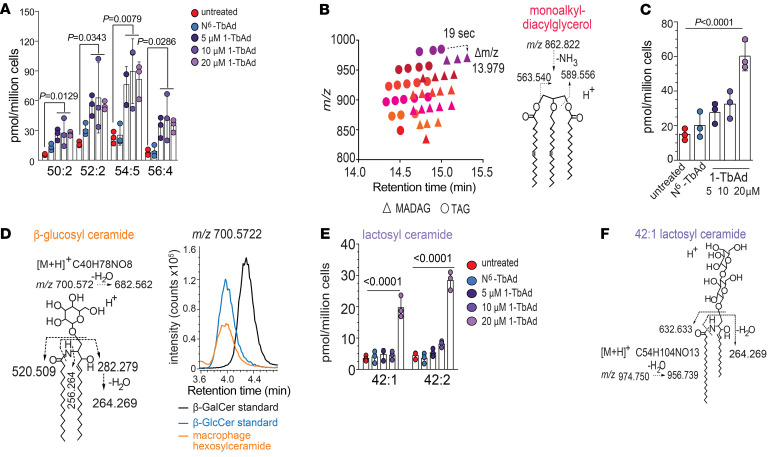
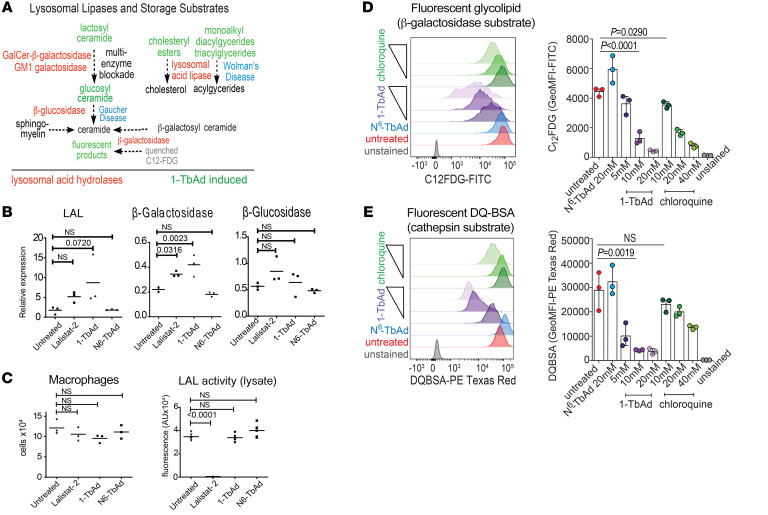
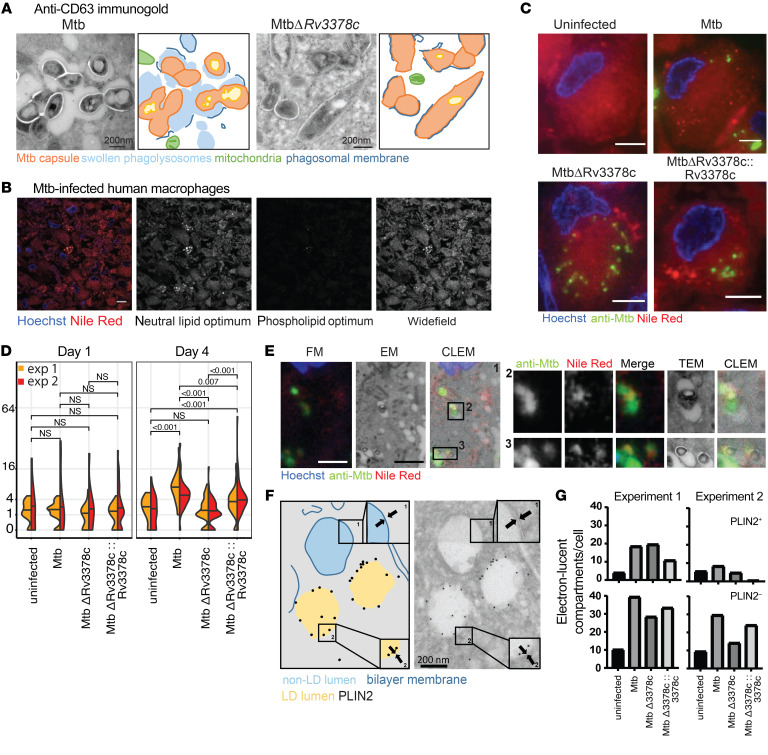
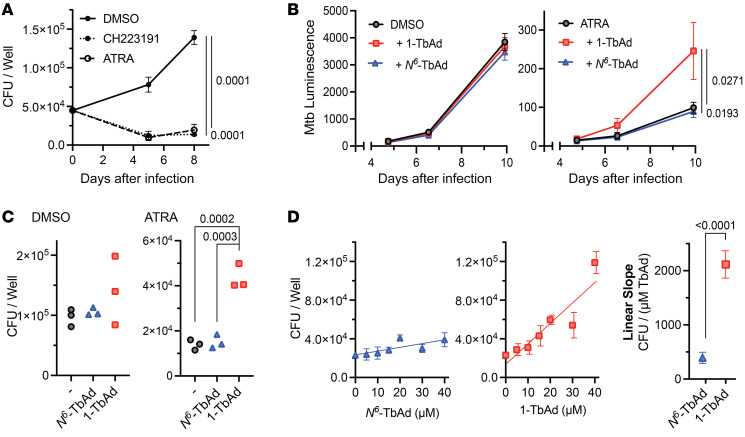
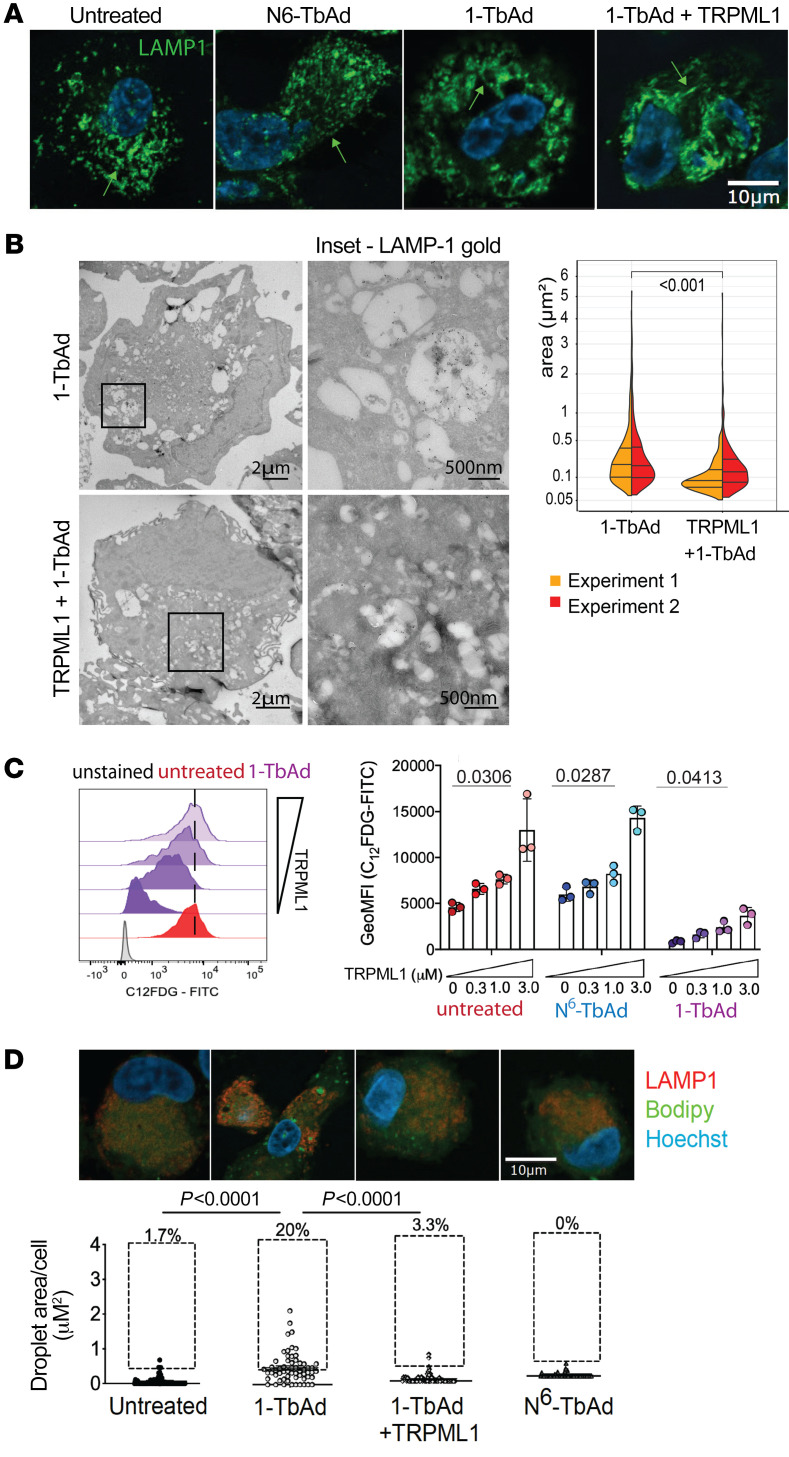
Induction of lipid-laden foamy macrophages is a cellular hallmark of tuberculosis (TB) disease, which involves the transformation of infected phagolysosomes from a site of killing into a nutrient-rich replicative niche. Here, we show that a terpenyl nucleoside shed from Mycobacterium tuberculosis, 1-tuberculosinyladenosine (1-TbAd), caused lysosomal maturation arrest and autophagy blockade, leading to lipid storage in M1 macrophages. Pure 1-TbAd, or infection with terpenyl nucleoside-producing M. tuberculosis, caused intralysosomal and peribacillary lipid storage patterns that matched both the molecules and subcellular locations known in foamy macrophages. Lipidomics showed that 1-TbAd induced storage of triacylglycerides and cholesterylesters and that 1-TbAd increased M. tuberculosis growth under conditions of restricted lipid access in macrophages. Furthermore, lipidomics identified 1-TbAd-induced lipid substrates that define Gaucher's disease, Wolman's disease, and other inborn lysosomal storage diseases. These data identify genetic and molecular causes of M. tuberculosis-induced lysosomal failure, leading to successful testing of an agonist of TRPML1 calcium channels that reverses lipid storage in cells. These data establish the host-directed cellular functions of an orphan effector molecule that promotes survival in macrophages, providing both an upstream cause and detailed picture of lysosome failure in foamy macrophages.
Keywords: Infectious disease; Macrophages; Microbiology; Tuberculosis.
Figures
Comment in
- The fat is in the lysosome: How Mycobacterium tuberculosis tricks macrophages into storing lipids doi: 10.1172/JCI168366
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
