

Organ Protection by Caloric Restriction Depends on Activation of the De Novo NAD+ Synthesis Pathway
- PMID: 36758124
- PMCID: PMC10125653
- DOI: 10.1681/ASN.0000000000000087
Organ Protection by Caloric Restriction Depends on Activation of the De Novo NAD+ Synthesis Pathway
Abstract
Significance statement: AKI is a major clinical complication leading to high mortality, but intensive research over the past decades has not led to targeted preventive or therapeutic measures. In rodent models, caloric restriction (CR) and transient hypoxia significantly prevent AKI and a recent comparative transcriptome analysis of murine kidneys identified kynureninase (KYNU) as a shared downstream target. The present work shows that KYNU strongly contributes to CR-mediated protection as a key player in the de novo nicotinamide adenine dinucleotide biosynthesis pathway. Importantly, the link between CR and NAD+ biosynthesis could be recapitulated in a human cohort.
Background: Clinical practice lacks strategies to treat AKI. Interestingly, preconditioning by hypoxia and caloric restriction (CR) is highly protective in rodent AKI models. However, the underlying molecular mechanisms of this process are unknown.
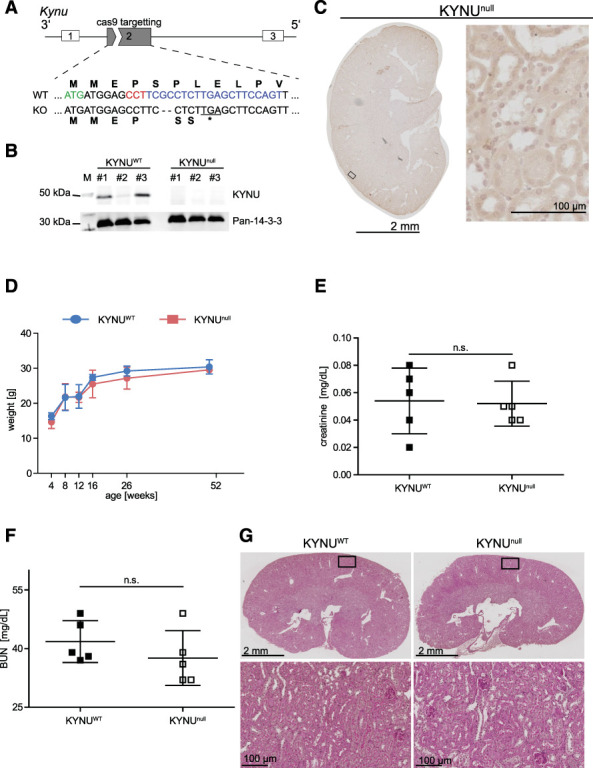
Methods: Kynureninase (KYNU) knockout mice were generated by Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) and comparative transcriptome, proteome and metabolite analyses of murine kidneys pre- and post-ischemia-reperfusion injury in the context of CR or ad libitum diet were performed. In addition, acetyl-lysin enrichment and mass spectrometry were used to assess protein acetylation.
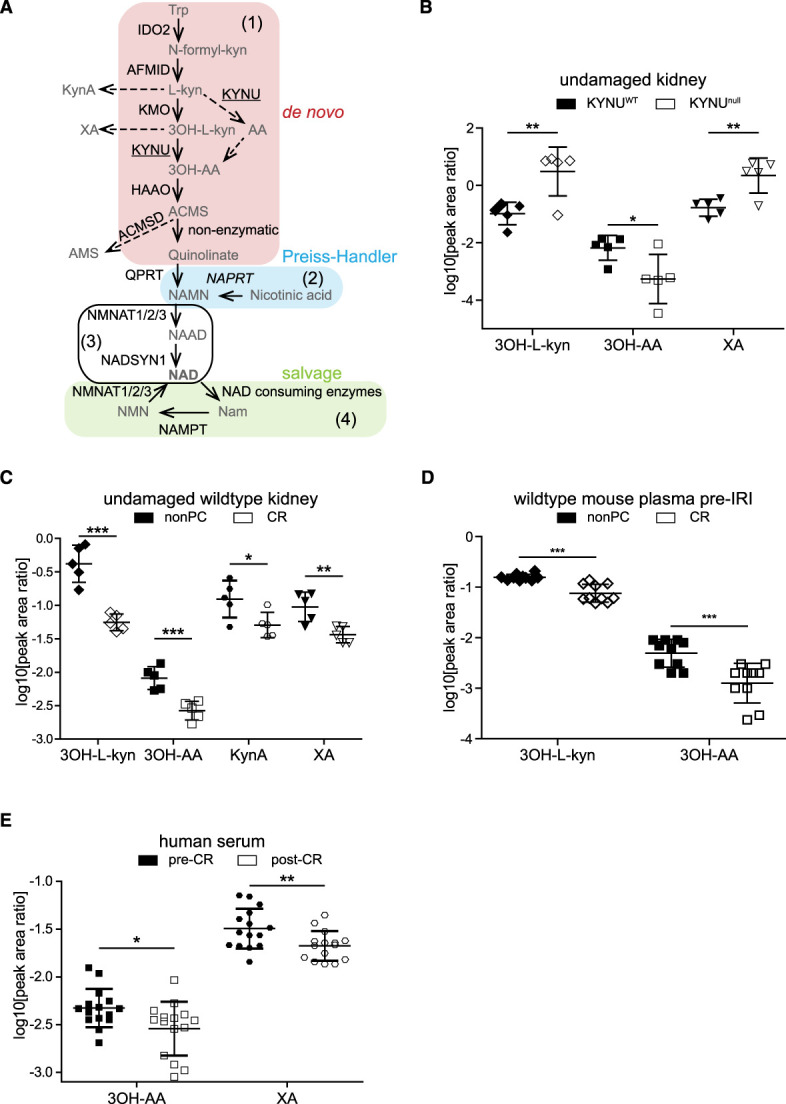
Results: We identified KYNU as a downstream target of CR and show that KYNU strongly contributes to the protective effect of CR. The KYNU-dependent de novo nicotinamide adenine dinucleotide (NAD+) biosynthesis pathway is necessary for CR-associated maintenance of NAD+ levels. This finding is associated with reduced protein acetylation in CR-treated animals, specifically affecting enzymes in energy metabolism. Importantly, the effect of CR on de novo NAD+ biosynthesis pathway metabolites can be recapitulated in humans.
Conclusions: CR induces the de novo NAD+ synthesis pathway in the context of IRI and is essential for its full nephroprotective potential. Differential protein acetylation may be the molecular mechanism underlying the relationship of NAD+, CR, and nephroprotection.
Copyright © 2023 by the American Society of Nephrology.
Conflict of interest statement
T. Benzing reports Consultancy: Advisory activity for Otsuka in the field of cystic kidney disease and hyponatremia; Honoraria: Speaker honoraria and travel support from Amgen, Hexal, Novartis, Otsuka, Roche, and Sanofi-Genzyme; and Speakers Bureau: Speaker honoraria and travel support from Amgen, Hexal, Novartis, Otsuka, Roche, and Sanofi-Genzyme. V. Burst reports Research Funding: Fresenius Kabi. M. Johnsen reports Honoraria: Heel. F.C. Koehler reports Consultancy: Atriva Therapeutics GmbH; and Research Funding: Else Kröner-Fresenius-Stiftung, German Research Foundation under Germany's Excellence Strategy—EXC 2030: CECAD–Excellent in Aging Research and Koeln Fortune program/Faculty of Medicine, University of Cologne, Germany. C. Lucas reports Employer: Merck Healthcare KgAA and Syft GmbH. R. Mueller reports Consultancy: Alnylam and Sanofi; Ownership Interest: Bayer, Chemocentryx, Novartis, Pfizer, Roche, and Santa Barbara Nutrients; Research Funding: Otsuka Pharmaceuticals and Thermo Fisher Scientific; all research funding was paid to the employer (Department II of Internal Medicine); Honoraria: Alnylam and Sanofi (consultancy agreements see above); and Advisory or Leadership Role: Board member of the WGIKD (ERA-EDTA); Editorial Board “
Figures





References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
