

Approximations of algorithmic and structural complexity validate cognitive-behavioral experimental results
- PMID: 36761393
- PMCID: PMC9904762
- DOI: 10.3389/fncom.2022.956074
Approximations of algorithmic and structural complexity validate cognitive-behavioral experimental results
Abstract
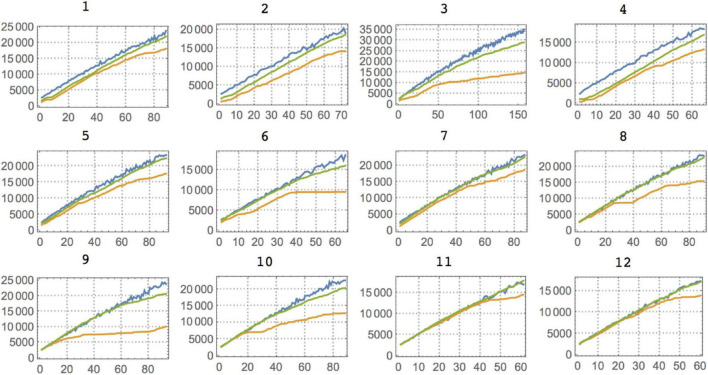
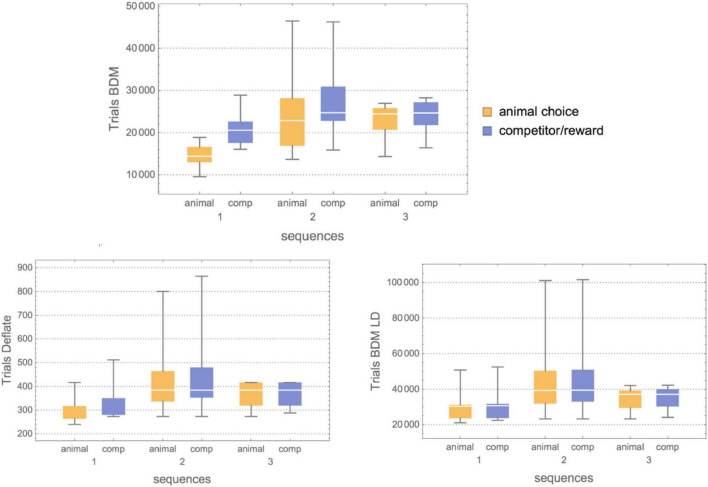
Being able to objectively characterize the intrinsic complexity of behavioral patterns resulting from human or animal decisions is fundamental for deconvolving cognition and designing autonomous artificial intelligence systems. Yet complexity is difficult in practice, particularly when strings are short. By numerically approximating algorithmic (Kolmogorov) complexity (K), we establish an objective tool to characterize behavioral complexity. Next, we approximate structural (Bennett's Logical Depth) complexity (LD) to assess the amount of computation required for generating a behavioral string. We apply our toolbox to three landmark studies of animal behavior of increasing sophistication and degree of environmental influence, including studies of foraging communication by ants, flight patterns of fruit flies, and tactical deception and competition (e.g., predator-prey) strategies. We find that ants harness the environmental condition in their internal decision process, modulating their behavioral complexity accordingly. Our analysis of flight (fruit flies) invalidated the common hypothesis that animals navigating in an environment devoid of stimuli adopt a random strategy. Fruit flies exposed to a featureless environment deviated the most from Levy flight, suggesting an algorithmic bias in their attempt to devise a useful (navigation) strategy. Similarly, a logical depth analysis of rats revealed that the structural complexity of the rat always ends up matching the structural complexity of the competitor, with the rats' behavior simulating algorithmic randomness. Finally, we discuss how experiments on how humans perceive randomness suggest the existence of an algorithmic bias in our reasoning and decision processes, in line with our analysis of the animal experiments. This contrasts with the view of the mind as performing faulty computations when presented with randomized items. In summary, our formal toolbox objectively characterizes external constraints on putative models of the "internal" decision process in humans and animals.
Keywords: Shannon Entropy; ant behavior; behavioral biases; behavioral sequences; communication complexity; tradeoffs of complexity measures.
Copyright © 2023 Zenil, Marshall and Tegnér.
Conflict of interest statement
HZ was employed by Oxford Immune Algorithmics Ltd. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
A Decomposition Method for Global Evaluation of Shannon Entropy and Local Estimations of Algorithmic Complexity.Entropy (Basel). 2018 Aug 15;20(8):605. doi: 10.3390/e20080605. Entropy (Basel). 2018. PMID: 33265694 Free PMC article.
-
Methods of information theory and algorithmic complexity for network biology.Semin Cell Dev Biol. 2016 Mar;51:32-43. doi: 10.1016/j.semcdb.2016.01.011. Epub 2016 Jan 21. Semin Cell Dev Biol. 2016. PMID: 26802516 Review.
-
Symmetry and Correspondence of Algorithmic Complexity over Geometric, Spatial and Topological Representations.Entropy (Basel). 2018 Jul 18;20(7):534. doi: 10.3390/e20070534. Entropy (Basel). 2018. PMID: 33265623 Free PMC article.
-
A Review of Graph and Network Complexity from an Algorithmic Information Perspective.Entropy (Basel). 2018 Jul 25;20(8):551. doi: 10.3390/e20080551. Entropy (Basel). 2018. PMID: 33265640 Free PMC article. Review.
-
Algorithmic complexity for short binary strings applied to psychology: a primer.Behav Res Methods. 2014 Sep;46(3):732-44. doi: 10.3758/s13428-013-0416-0. Behav Res Methods. 2014. PMID: 24311059
Cited by
-
A Review of Methods for Estimating Algorithmic Complexity: Options, Challenges, and New Directions.Entropy (Basel). 2020 May 30;22(6):612. doi: 10.3390/e22060612. Entropy (Basel). 2020. PMID: 33286384 Free PMC article. Review.
-
Information Theory Opens New Dimensions in Experimental Studies of Animal Behaviour and Communication.Animals (Basel). 2023 Mar 26;13(7):1174. doi: 10.3390/ani13071174. Animals (Basel). 2023. PMID: 37048430 Free PMC article. Review.
-
Inform: Efficient Information-Theoretic Analysis of Collective Behaviors.Front Robot AI. 2018 Jun 11;5:60. doi: 10.3389/frobt.2018.00060. eCollection 2018. Front Robot AI. 2018. PMID: 33718436 Free PMC article.
-
Algorithmic Entropy and Landauer's Principle Link Microscopic System Behaviour to the Thermodynamic Entropy.Entropy (Basel). 2018 Oct 17;20(10):798. doi: 10.3390/e20100798. Entropy (Basel). 2018. PMID: 33265885 Free PMC article.
References
-
- Bennett C. (1988). “Logical depth and physical complexity,” in The universal turing machine–a half-century survey, ed. Herken R. (Oxford: Oxford University Press; ), 227–257.
-
- Chaitin G. (1969). On the length of programs for computing finite binary sequences: Statistical considerations. J. ACM 16 145–159. 10.1145/321495.321506 - DOI
LinkOut - more resources
Full Text Sources
Research Materials