

Ribosome biogenesis factors-from names to functions
- PMID: 36762427
- PMCID: PMC10068337
- DOI: 10.15252/embj.2022112699
Ribosome biogenesis factors-from names to functions
Abstract
The assembly of ribosomal subunits is a highly orchestrated process that involves a huge cohort of accessory factors. Most eukaryotic ribosome biogenesis factors were first identified by genetic screens and proteomic approaches of pre-ribosomal particles in Saccharomyces cerevisiae. Later, research on human ribosome synthesis not only demonstrated that the requirement for many of these factors is conserved in evolution, but also revealed the involvement of additional players, reflecting a more complex assembly pathway in mammalian cells. Yet, it remained a challenge for the field to assign a function to many of the identified factors and to reveal their molecular mode of action. Over the past decade, structural, biochemical, and cellular studies have largely filled this gap in knowledge and led to a detailed understanding of the molecular role that many of the players have during the stepwise process of ribosome maturation. Such detailed knowledge of the function of ribosome biogenesis factors will be key to further understand and better treat diseases linked to disturbed ribosome assembly, including ribosomopathies, as well as different types of cancer.
Keywords: pre-rRNA processing; pre-ribosomal particle; ribosomal subunit; ribosome biogenesis factor; ribosome synthesis.
© 2023 The Authors. Published under the terms of the CC BY NC ND 4.0 license.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
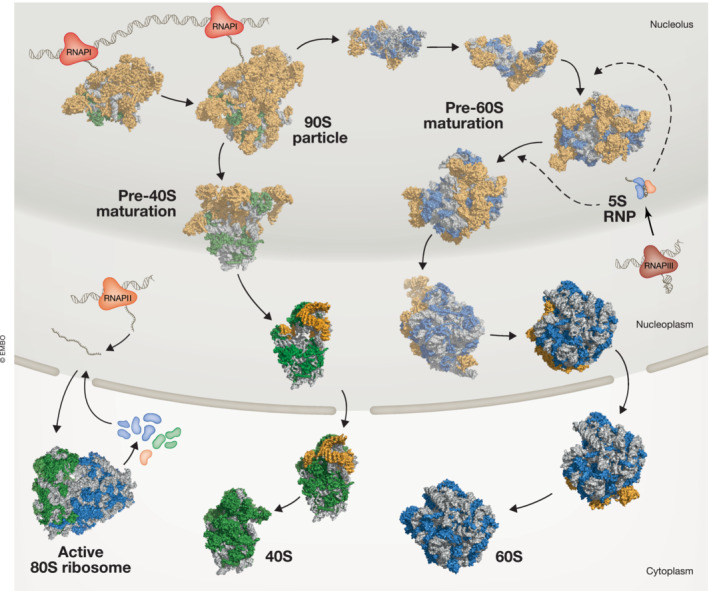
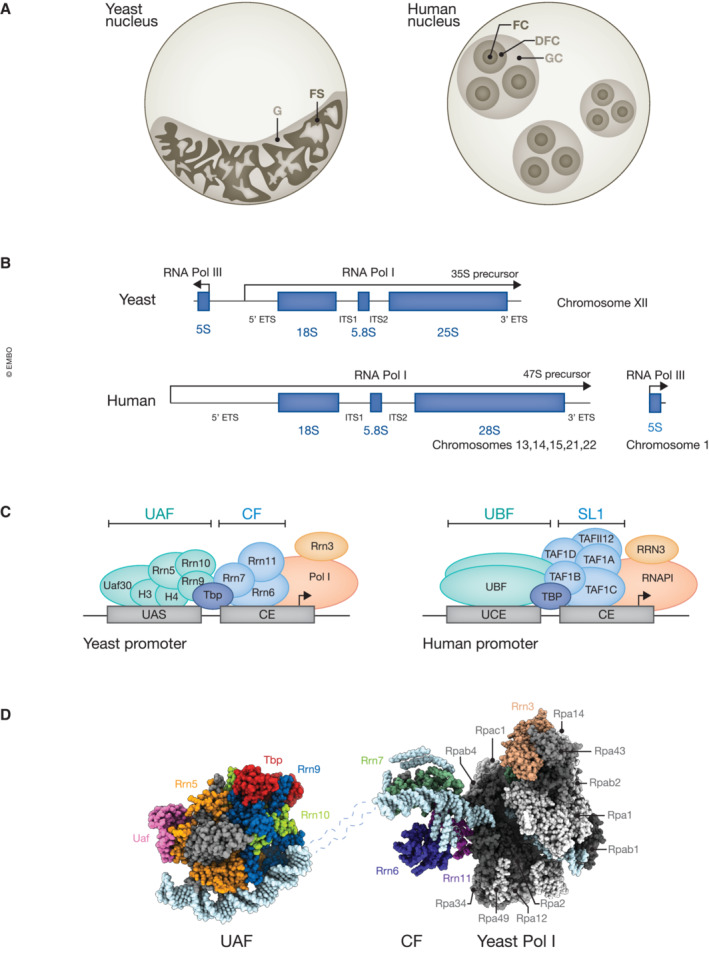
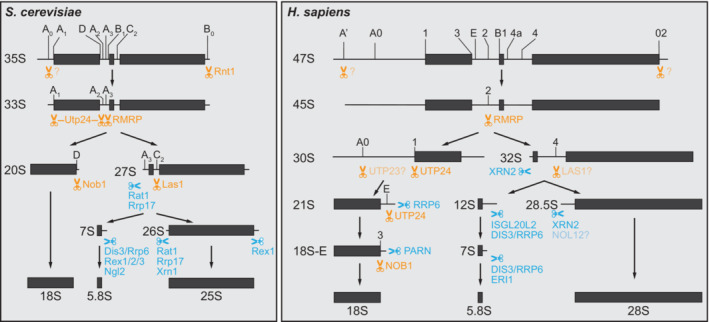
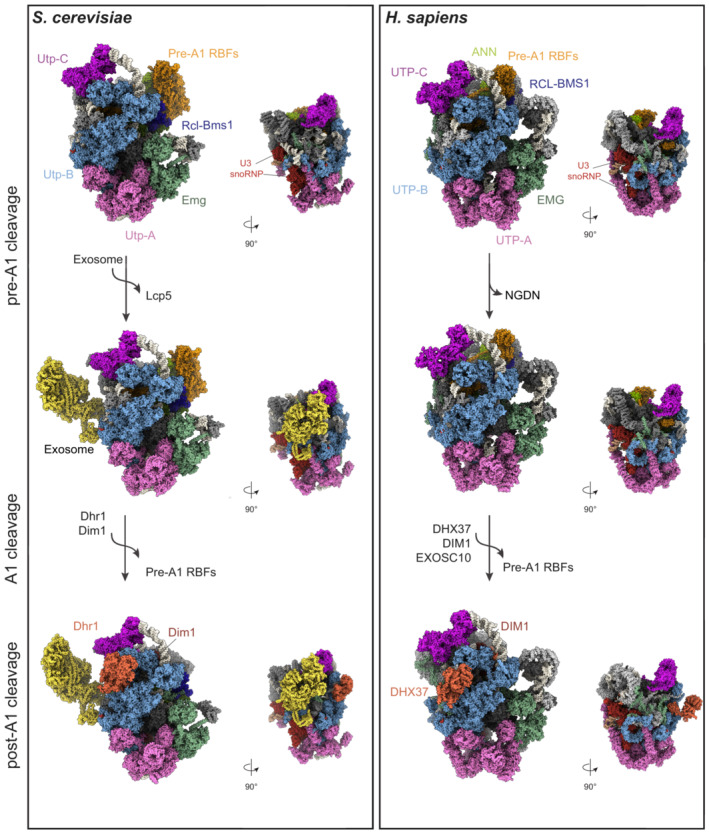
Similar articles
-
Coevolution of ribosomal RNA expansion segment 7L and assembly factor Noc2p specializes the ribosome biogenesis pathway between Saccharomyces cerevisiae and Candida albicans.Nucleic Acids Res. 2021 May 7;49(8):4655-4667. doi: 10.1093/nar/gkab218. Nucleic Acids Res. 2021. PMID: 33823547 Free PMC article.
-
Role of the yeast ribosomal protein L16 in ribosome biogenesis.FEBS J. 2016 Aug;283(16):2968-85. doi: 10.1111/febs.13797. Epub 2016 Jul 15. FEBS J. 2016. PMID: 27374275
-
Las1 interacts with Grc3 polynucleotide kinase and is required for ribosome synthesis in Saccharomyces cerevisiae.Nucleic Acids Res. 2013 Jan;41(2):1135-50. doi: 10.1093/nar/gks1086. Epub 2012 Nov 21. Nucleic Acids Res. 2013. PMID: 23175604 Free PMC article.
-
Ribosome assembly in eukaryotes.Gene. 2003 Aug 14;313:17-42. doi: 10.1016/s0378-1119(03)00629-2. Gene. 2003. PMID: 12957375 Review.
-
RNA folding and functions of RNA helicases in ribosome biogenesis.RNA Biol. 2022 Jan;19(1):781-810. doi: 10.1080/15476286.2022.2079890. RNA Biol. 2022. PMID: 35678541 Free PMC article. Review.
Cited by
-
Experimental upregulation of developmentally downregulated ribosomal protein large subunits 7 and 7A promotes axon regeneration after injury in vivo.Exp Neurol. 2023 Oct;368:114510. doi: 10.1016/j.expneurol.2023.114510. Epub 2023 Aug 24. Exp Neurol. 2023. PMID: 37633482 Free PMC article.
-
Multi-omics profiling reveals dysregulated ribosome biogenesis and impaired cell proliferation following knockout of CDR2L.BMC Cancer. 2024 May 27;24(1):645. doi: 10.1186/s12885-024-12399-z. BMC Cancer. 2024. PMID: 38802745 Free PMC article.
-
USP36 SUMOylates Las1L and Promotes Its Function in Pre-Ribosomal RNA ITS2 Processing.Cancer Res Commun. 2024 Oct 1;4(10):2835-2845. doi: 10.1158/2767-9764.CRC-24-0312. Cancer Res Commun. 2024. PMID: 39356143 Free PMC article.
-
Critical and differential roles of eIF4A1 and eIF4A2 in B-cell development and function.Cell Mol Immunol. 2025 Jan;22(1):40-53. doi: 10.1038/s41423-024-01234-x. Epub 2024 Nov 8. Cell Mol Immunol. 2025. PMID: 39516355 Free PMC article.
-
From Flies to Humans: A Conserved Role of CEBPZ, NOC2L, and NOC3L in rRNA Processing and Tumorigenesis.bioRxiv [Preprint]. 2025 Apr 23:2025.01.11.632529. doi: 10.1101/2025.01.11.632529. bioRxiv. 2025. Update in: J Cell Sci. 2025 Aug 19:jcs.264096. doi: 10.1242/jcs.264096. PMID: 39868138 Free PMC article. Updated. Preprint.
References
-
- Allmang C, Tollervey D (1998) The role of the 3′ external transcribed spacer in yeast pre‐rRNA processing. J Mol Biol 278: 67–78 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
