

A riboswitch separated from its ribosome-binding site still regulates translation
- PMID: 36762498
- PMCID: PMC10018353
- DOI: 10.1093/nar/gkad056
A riboswitch separated from its ribosome-binding site still regulates translation
Abstract
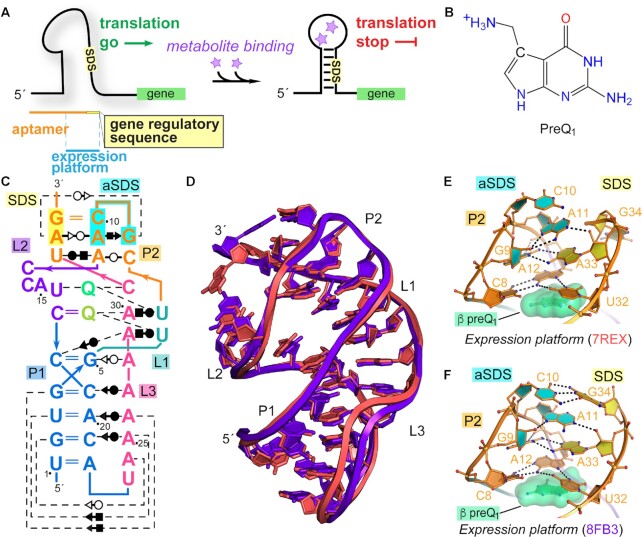
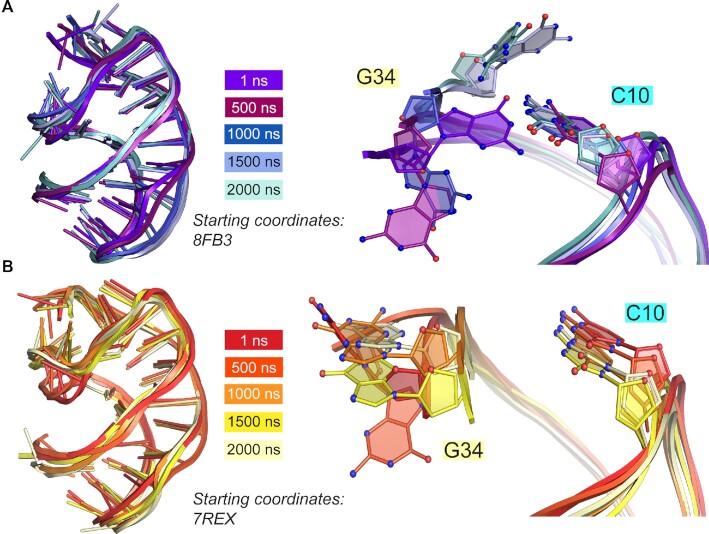
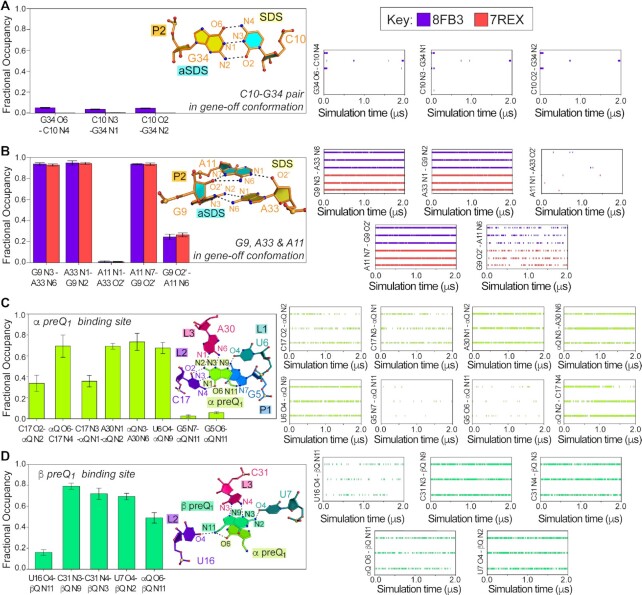
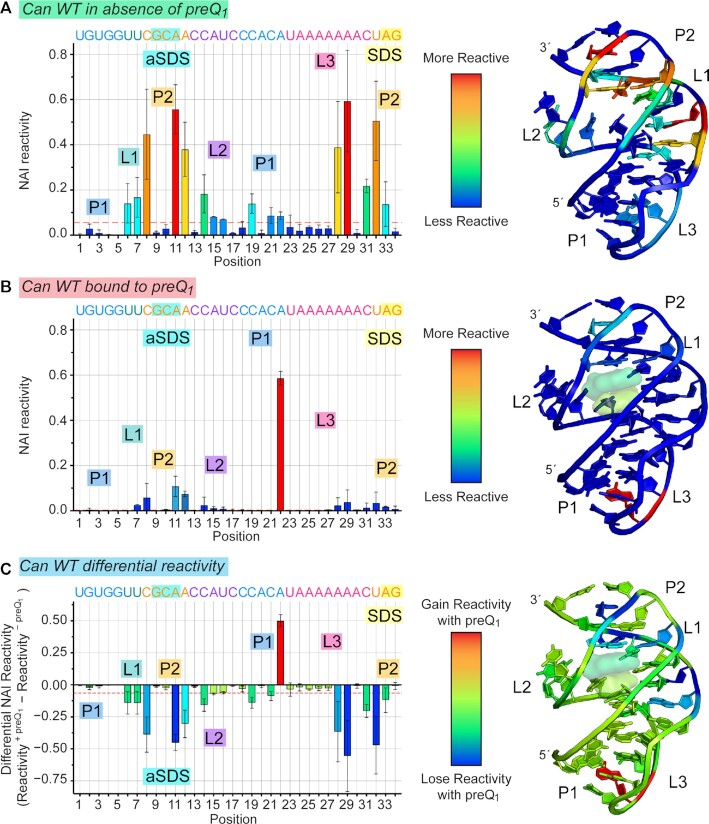
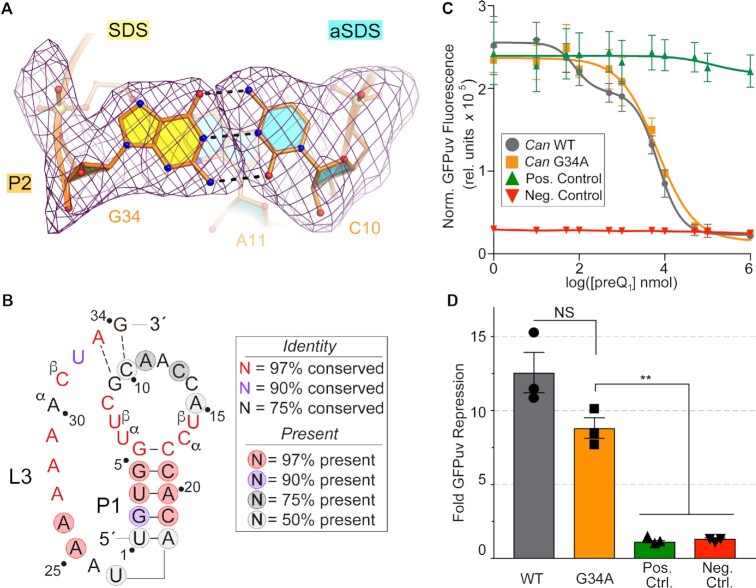
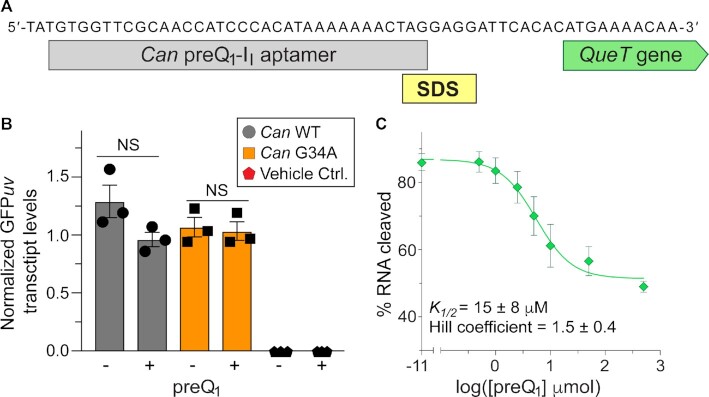
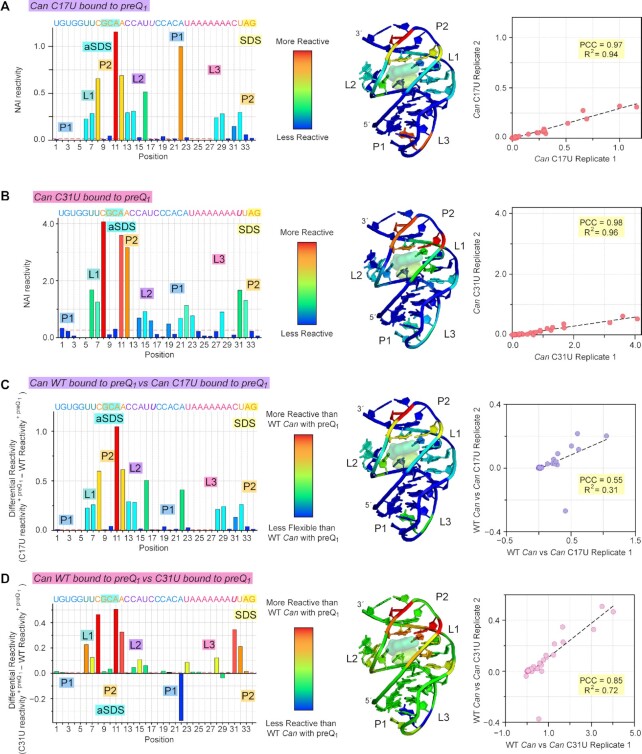
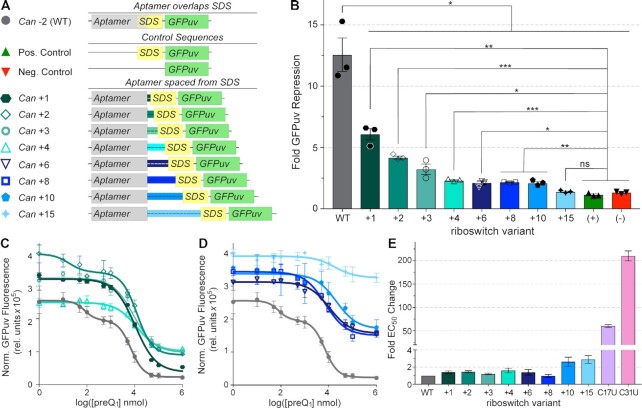
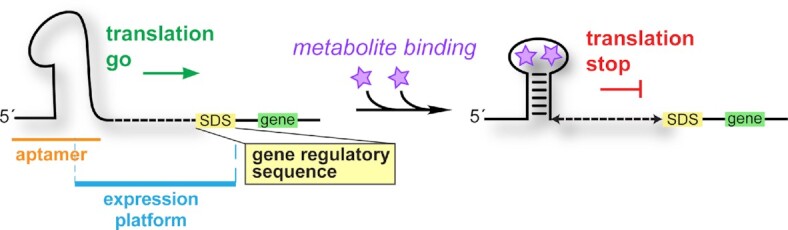
Riboswitches regulate downstream gene expression by binding cellular metabolites. Regulation of translation initiation by riboswitches is posited to occur by metabolite-mediated sequestration of the Shine-Dalgarno sequence (SDS), causing bypass by the ribosome. Recently, we solved a co-crystal structure of a prequeuosine1-sensing riboswitch from Carnobacterium antarcticum that binds two metabolites in a single pocket. The structure revealed that the second nucleotide within the gene-regulatory SDS, G34, engages in a crystal contact, obscuring the molecular basis of gene regulation. Here, we report a co-crystal structure wherein C10 pairs with G34. However, molecular dynamics simulations reveal quick dissolution of the pair, which fails to reform. Functional and chemical probing assays inside live bacterial cells corroborate the dispensability of the C10-G34 pair in gene regulation, leading to the hypothesis that the compact pseudoknot fold is sufficient for translation attenuation. Remarkably, the C. antarcticum aptamer retained significant gene-regulatory activity when uncoupled from the SDS using unstructured spacers up to 10 nucleotides away from the riboswitch-akin to steric-blocking employed by sRNAs. Accordingly, our work reveals that the RNA fold regulates translation without SDS sequestration, expanding known riboswitch-mediated gene-regulatory mechanisms. The results infer that riboswitches exist wherein the SDS is not embedded inside a stable fold.
© The Author(s) 2023. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Similar articles
-
Structure and function analysis of a type III preQ1-I riboswitch from Escherichia coli reveals direct metabolite sensing by the Shine-Dalgarno sequence.J Biol Chem. 2023 Oct;299(10):105208. doi: 10.1016/j.jbc.2023.105208. Epub 2023 Sep 1. J Biol Chem. 2023. PMID: 37660906 Free PMC article.
-
Knotty is nice: Metabolite binding and RNA-mediated gene regulation by the preQ1 riboswitch family.J Biol Chem. 2024 Dec;300(12):107951. doi: 10.1016/j.jbc.2024.107951. Epub 2024 Oct 30. J Biol Chem. 2024. PMID: 39486689 Free PMC article. Review.
-
Structural analysis of a class III preQ1 riboswitch reveals an aptamer distant from a ribosome-binding site regulated by fast dynamics.Proc Natl Acad Sci U S A. 2015 Jul 7;112(27):E3485-94. doi: 10.1073/pnas.1503955112. Epub 2015 Jun 23. Proc Natl Acad Sci U S A. 2015. PMID: 26106162 Free PMC article.
-
Nucleobase mutants of a bacterial preQ1-II riboswitch that uncouple metabolite sensing from gene regulation.J Biol Chem. 2020 Feb 28;295(9):2555-2567. doi: 10.1074/jbc.RA119.010755. Epub 2019 Oct 28. J Biol Chem. 2020. PMID: 31659117 Free PMC article.
-
[The adenine riboswitch: a new gene regulation mechanism].Med Sci (Paris). 2006 Dec;22(12):1053-9. doi: 10.1051/medsci/200622121053. Med Sci (Paris). 2006. PMID: 17156726 Review. French.
Cited by
-
Biosynthesis and function of 7-deazaguanine derivatives in bacteria and phages.Microbiol Mol Biol Rev. 2024 Mar 27;88(1):e0019923. doi: 10.1128/mmbr.00199-23. Epub 2024 Feb 29. Microbiol Mol Biol Rev. 2024. PMID: 38421302 Free PMC article. Review.
-
Applications and Tuning Strategies for Transcription Factor-Based Metabolite Biosensors.Biosensors (Basel). 2023 Mar 28;13(4):428. doi: 10.3390/bios13040428. Biosensors (Basel). 2023. PMID: 37185503 Free PMC article. Review.
-
Comparative analysis of RNA 3D structure prediction methods: towards enhanced modeling of RNA-ligand interactions.Nucleic Acids Res. 2024 Jul 22;52(13):7465-7486. doi: 10.1093/nar/gkae541. Nucleic Acids Res. 2024. PMID: 38917327 Free PMC article.
-
Electrostatic Anchoring in RNA-Ligand Design─Dissecting the Effects of Positive Charges on Affinity, Selectivity, Binding Kinetics, and Thermodynamics.J Med Chem. 2025 Apr 24;68(8):8659-8678. doi: 10.1021/acs.jmedchem.5c00339. Epub 2025 Apr 7. J Med Chem. 2025. PMID: 40191889 Free PMC article.
-
Structure and function analysis of a type III preQ1-I riboswitch from Escherichia coli reveals direct metabolite sensing by the Shine-Dalgarno sequence.J Biol Chem. 2023 Oct;299(10):105208. doi: 10.1016/j.jbc.2023.105208. Epub 2023 Sep 1. J Biol Chem. 2023. PMID: 37660906 Free PMC article.
References
-
- Nahvi A., Sudarsan N., Ebert M.S., Zou X., Brown K.L., Breaker R.R.. Genetic control by a metabolite binding mRNA. Chem. Biol. 2002; 9:1043. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
