

Pericytes and the Control of Blood Flow in Brain and Heart
- PMID: 36763972
- PMCID: PMC10280497
- DOI: 10.1146/annurev-physiol-031522-034807
Pericytes and the Control of Blood Flow in Brain and Heart
Abstract
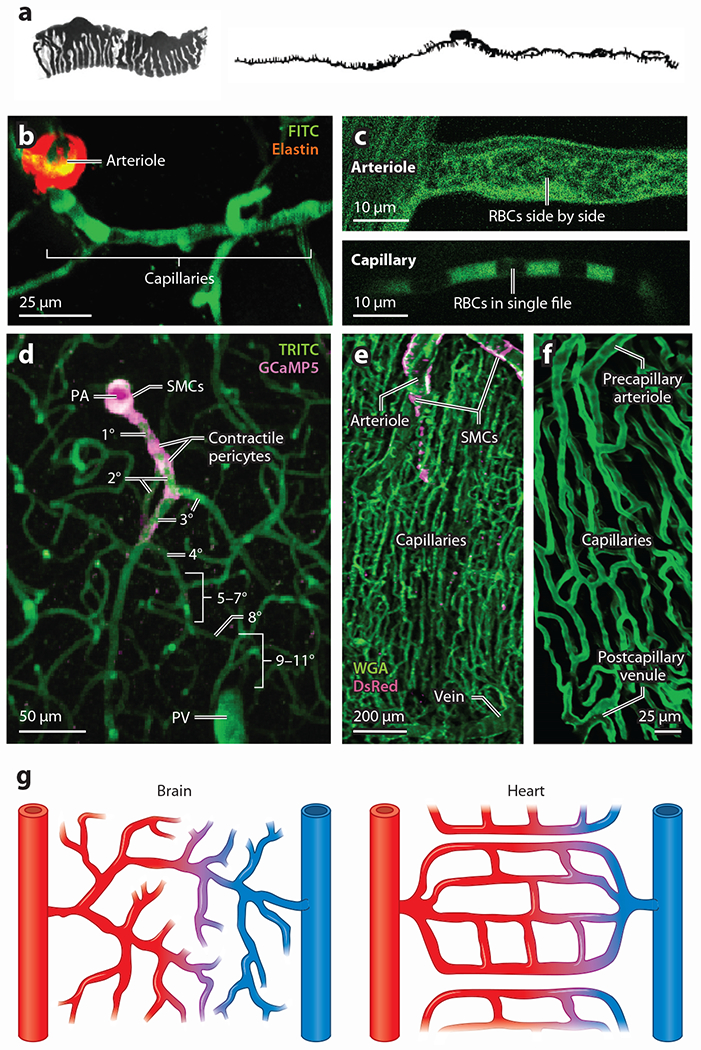
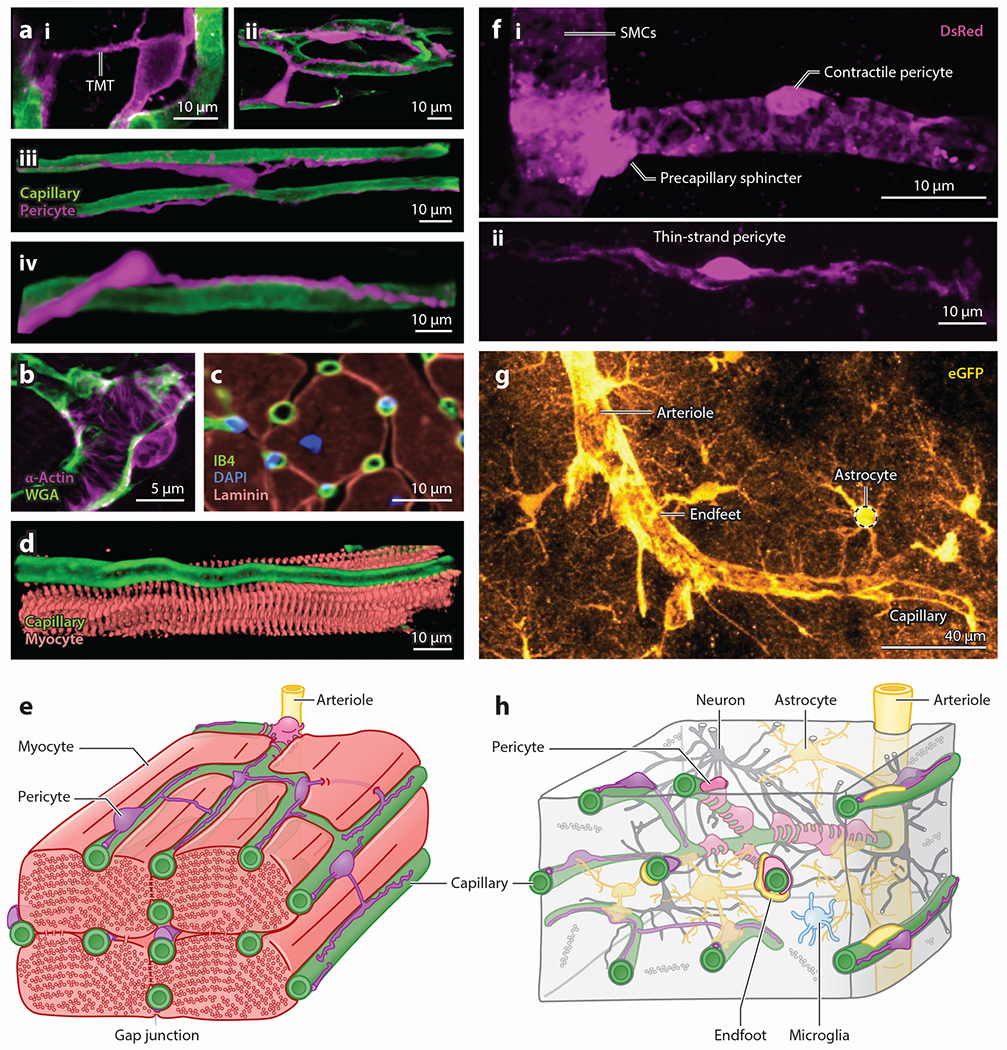
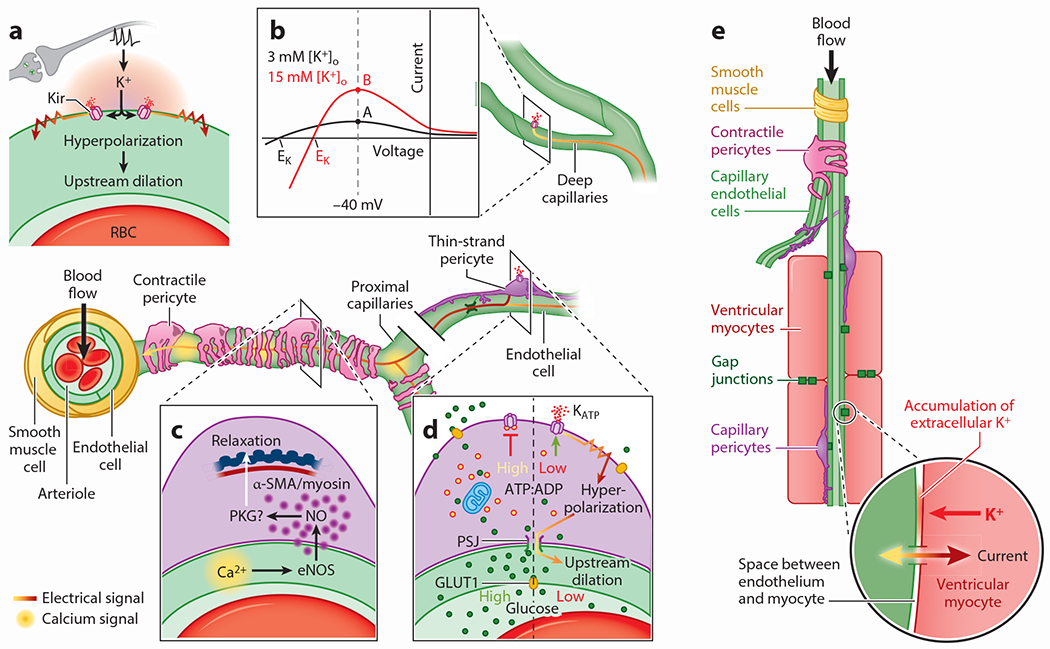
Pericytes, attached to the surface of capillaries, play an important role in regulating local blood flow. Using optogenetic tools and genetically encoded reporters in conjunction with confocal and multiphoton imaging techniques, the 3D structure, anatomical organization, and physiology of pericytes have recently been the subject of detailed examination. This work has revealed novel functions of pericytes and morphological features such as tunneling nanotubes in brain and tunneling microtubes in heart. Here, we discuss the state of our current understanding of the roles of pericytes in blood flow control in brain and heart, where functions may differ due to the distinct spatiotemporal metabolic requirements of these tissues. We also outline the novel concept of electro-metabolic signaling, a universal mechanistic framework that links tissue metabolic state with blood flow regulation by pericytes and vascular smooth muscle cells, with capillary KATP and Kir2.1 channels as primary sensors. Finally, we present major unresolved questions and outline how they can be addressed.
Keywords: brain; calcium; electro-metabolic signaling; heart; ion channels; metabolism; neurovascular coupling; pericytes; potassium.
Figures
References
-
- Eberth C. 1871. Handbuch der Lehre von der Gewegen des Menschen und der Tiere. Vol. 1. Leipzig, Ger.: Engelmann
-
- Rouget C. 1873. Mémoire sur le développement, la structure et les proprietés physiologiques des capillaires sanguins et lymphatiques. Arch. Physiol. Norm. Pathol 5:603–63
-
- Zimmermann KW. 1923. Der feinere Bau der Blutcapillaren. Z. Anat. Entwicklungsgesch 68(1):29–109
-
- Krogh A. 1920. A contribution to the physiology of the capillaries. Nobel Lecture, Dec. 11. https://www.nobelprize.org/prizes/medicine/1920/krogh/lecture/
-
- Feigl EO. 1983. Coronary physiology. Physiol. Rev 63(1):1–205 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
