

Barrier properties of Nup98 FG phases ruled by FG motif identity and inter-FG spacer length
- PMID: 36765044
- PMCID: PMC9918544
- DOI: 10.1038/s41467-023-36331-4
Barrier properties of Nup98 FG phases ruled by FG motif identity and inter-FG spacer length
Abstract
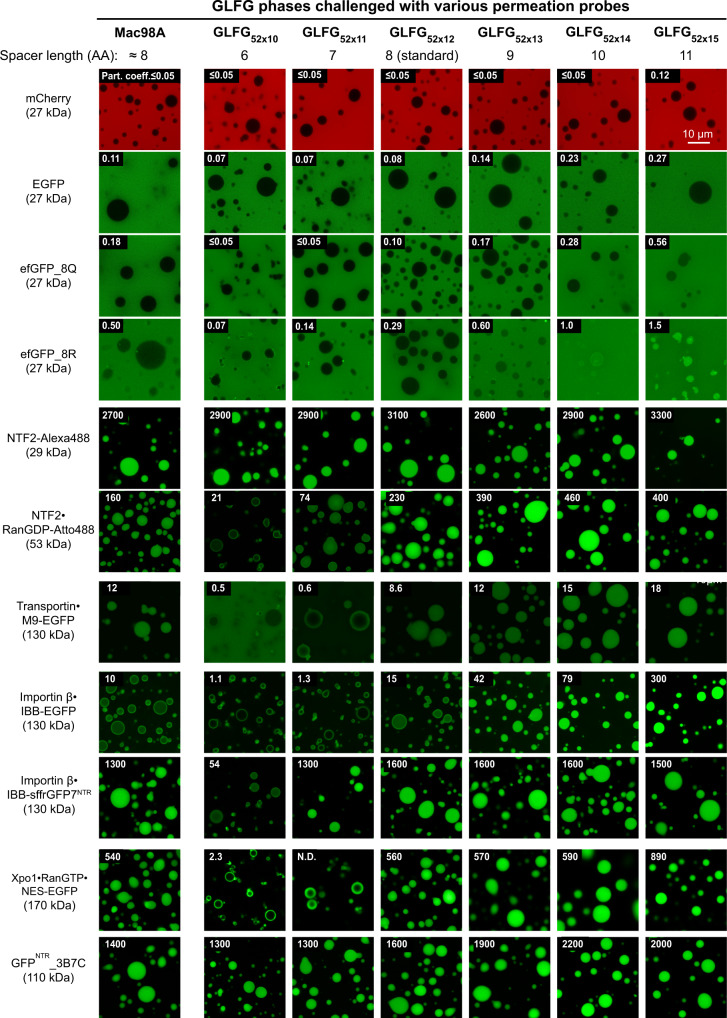
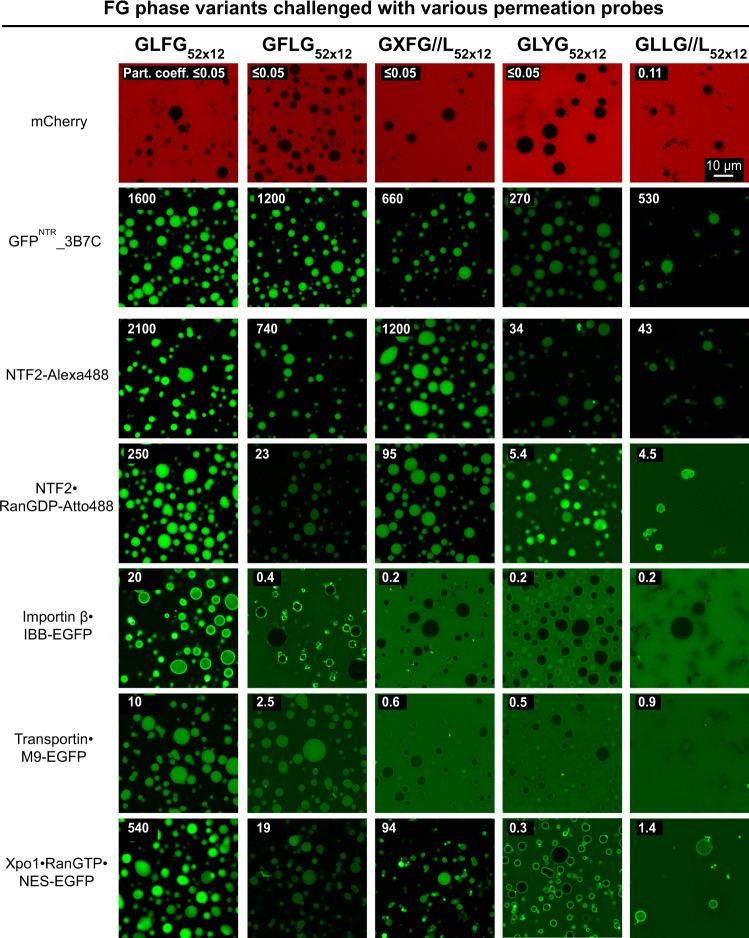
Nup98 FG repeat domains comprise hydrophobic FG motifs linked through uncharged spacers. FG motifs capture nuclear transport receptors (NTRs) during nuclear pore complex (NPC) passage, confer inter-repeat cohesion, and condense the domains into a selective phase with NPC-typical barrier properties. We show that shortening inter-FG spacers enhances cohesion, increases phase density, and tightens such barrier - all consistent with a sieve-like phase. Phase separation tolerates mutating the Nup98-typical GLFG motifs, provided domain-hydrophobicity remains preserved. NTR-entry, however, is sensitive to (certain) deviations from canonical FG motifs, suggesting co-evolutionary adaptation. Unexpectedly, we observed that arginines promote FG-phase-entry apparently also by hydrophobic interactions/ hydrogen-bonding and not just through cation-π interactions. Although incompatible with NTR·cargo complexes, a YG phase displays remarkable transport selectivity, particularly for engineered GFPNTR-variants. GLFG to FSFG mutations make the FG phase hypercohesive, precluding NTR-entry. Extending spacers relaxes this hypercohesion. Thus, antagonism between cohesion and NTR·FG interactions is key to transport selectivity.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures








Similar articles
-
Nup98 FG domains from diverse species spontaneously phase-separate into particles with nuclear pore-like permselectivity.Elife. 2015 Jan 6;4:e04251. doi: 10.7554/eLife.04251. Elife. 2015. PMID: 25562883 Free PMC article.
-
Surface Properties Determining Passage Rates of Proteins through Nuclear Pores.Cell. 2018 Jun 28;174(1):202-217.e9. doi: 10.1016/j.cell.2018.05.045. Cell. 2018. PMID: 29958108
-
Systematic analysis of barrier-forming FG hydrogels from Xenopus nuclear pore complexes.EMBO J. 2013 Jan 23;32(2):204-18. doi: 10.1038/emboj.2012.302. Epub 2012 Nov 30. EMBO J. 2013. PMID: 23202855 Free PMC article.
-
Biomechanics of the transport barrier in the nuclear pore complex.Semin Cell Dev Biol. 2017 Aug;68:42-51. doi: 10.1016/j.semcdb.2017.05.007. Epub 2017 May 12. Semin Cell Dev Biol. 2017. PMID: 28506890 Review.
-
The selective permeability barrier in the nuclear pore complex.Nucleus. 2016 Sep 2;7(5):430-446. doi: 10.1080/19491034.2016.1238997. Epub 2016 Sep 27. Nucleus. 2016. PMID: 27673359 Free PMC article. Review.
Cited by
-
Channel width modulates the permeability of DNA origami based nuclear pore mimics.bioRxiv [Preprint]. 2024 May 12:2024.05.09.593438. doi: 10.1101/2024.05.09.593438. bioRxiv. 2024. Update in: Sci Adv. 2024 Nov 15;10(46):eadq8773. doi: 10.1126/sciadv.adq8773. PMID: 38766144 Free PMC article. Updated. Preprint.
-
Nuclear pore and nucleocytoplasmic transport impairment in oxidative stress-induced neurodegeneration: relevance to molecular mechanisms in Pathogenesis of Parkinson's and other related neurodegenerative diseases.Mol Neurodegener. 2024 Nov 23;19(1):87. doi: 10.1186/s13024-024-00774-0. Mol Neurodegener. 2024. PMID: 39578912 Free PMC article. Review.
-
Visualizing the disordered nuclear transport machinery in situ.Nature. 2023 May;617(7959):162-169. doi: 10.1038/s41586-023-05990-0. Epub 2023 Apr 26. Nature. 2023. PMID: 37100914 Free PMC article.
-
Molecular Crowding: Physiologic Sensing and Control.Annu Rev Physiol. 2024 Feb 12;86:429-452. doi: 10.1146/annurev-physiol-042222-025920. Epub 2023 Nov 6. Annu Rev Physiol. 2024. PMID: 37931170 Free PMC article. Review.
-
Nanoscale interactions of arginine-containing dipeptide repeats with nuclear pore complexes as measured by transient scanning electrochemical microscopy.Chem Sci. 2024 Aug 30;15(38):15639-46. doi: 10.1039/d4sc05063k. Online ahead of print. Chem Sci. 2024. PMID: 39246336 Free PMC article.
References
-
- Hampoelz B, Andres-Pons A, Kastritis P, Beck M. Structure and assembly of the nuclear pore complex. Annu. Rev. Biophys. 2019;48:515–536. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
