

Hedgehog is relayed through dynamic heparan sulfate interactions to shape its gradient
- PMID: 36765094
- PMCID: PMC9918555
- DOI: 10.1038/s41467-023-36450-y
Hedgehog is relayed through dynamic heparan sulfate interactions to shape its gradient
Abstract
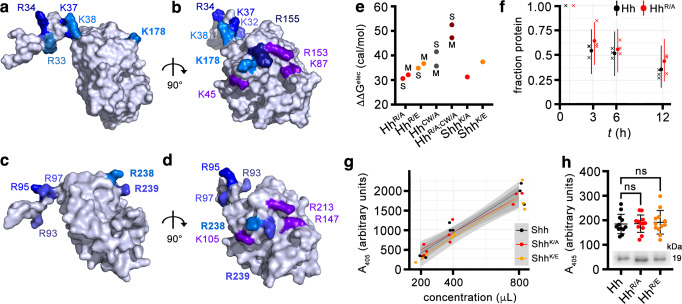
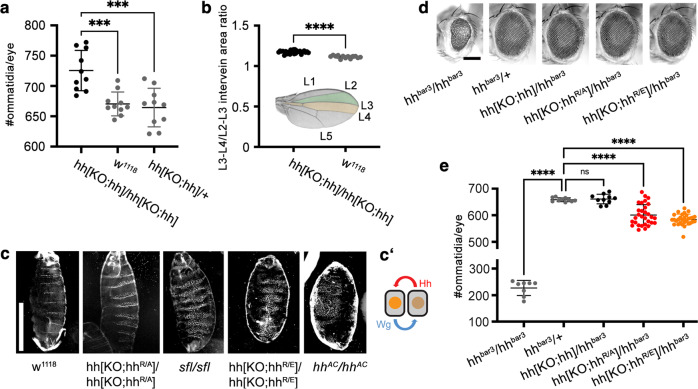
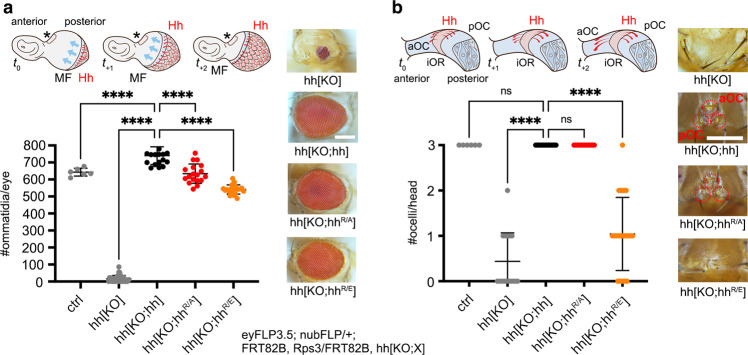
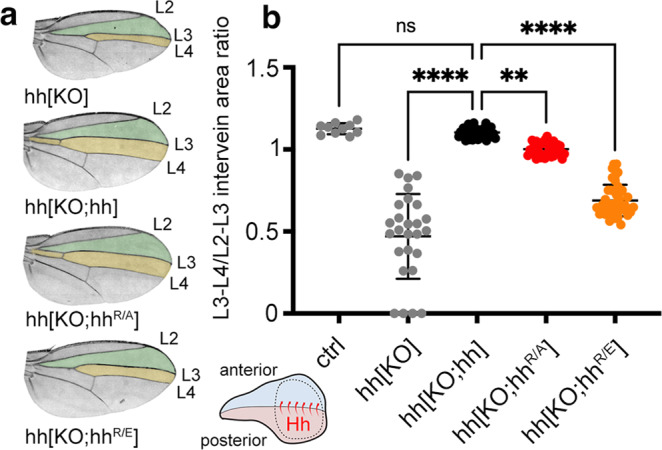
Cellular differentiation is directly determined by concentration gradients of morphogens. As a central model for gradient formation during development, Hedgehog (Hh) morphogens spread away from their source to direct growth and pattern formation in Drosophila wing and eye discs. What is not known is how extracellular Hh spread is achieved and how it translates into precise gradients. Here we show that two separate binding areas located on opposite sides of the Hh molecule can interact directly and simultaneously with two heparan sulfate (HS) chains to temporarily cross-link the chains. Mutated Hh lacking one fully functional binding site still binds HS but shows reduced HS cross-linking. This, in turn, impairs Hhs ability to switch between both chains in vitro and results in striking Hh gradient hypomorphs in vivo. The speed and propensity of direct Hh switching between HS therefore shapes the Hh gradient, revealing a scalable design principle in morphogen-patterned tissues.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
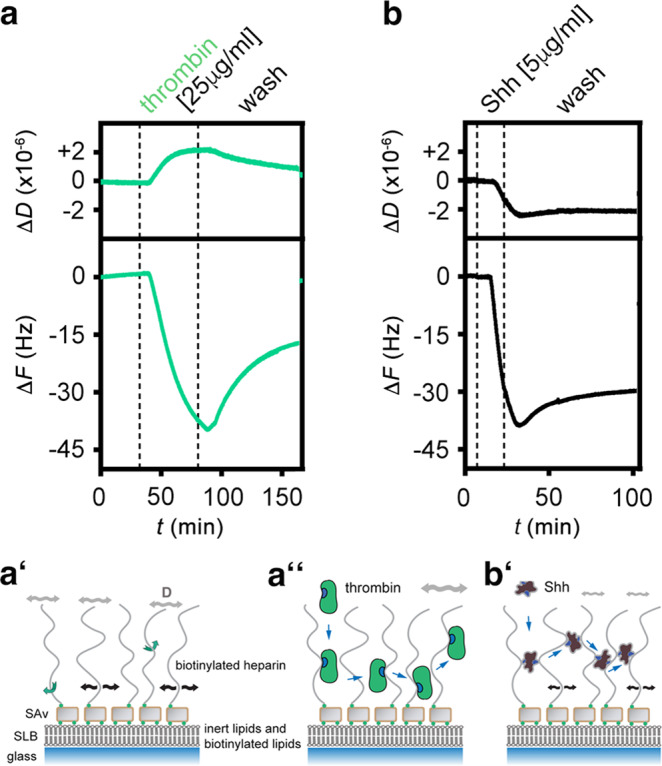
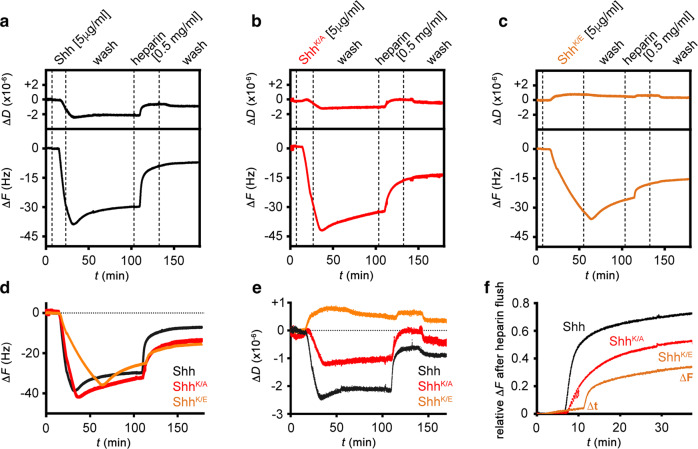
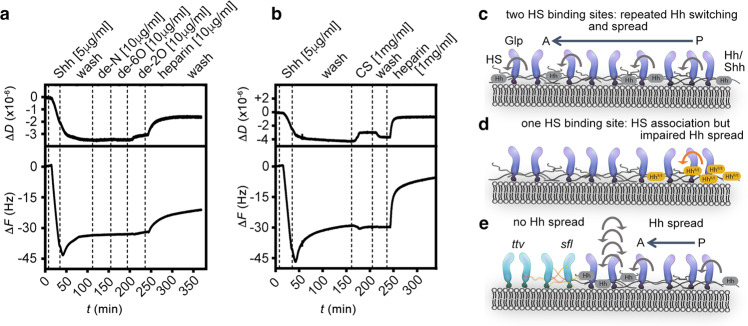
Similar articles
-
Drosophila hedgehog signaling range and robustness depend on direct and sustained heparan sulfate interactions.Front Mol Biosci. 2023 Feb 22;10:1130064. doi: 10.3389/fmolb.2023.1130064. eCollection 2023. Front Mol Biosci. 2023. PMID: 36911531 Free PMC article.
-
Hedgehog on the move: a precise spatial control of Hedgehog dispersion shapes the gradient.Curr Opin Genet Dev. 2013 Aug;23(4):363-73. doi: 10.1016/j.gde.2013.04.011. Epub 2013 Jun 5. Curr Opin Genet Dev. 2013. PMID: 23747033 Review.
-
Drosophila hedgehog can act as a morphogen in the absence of regulated Ci processing.Elife. 2020 Oct 21;9:e61083. doi: 10.7554/eLife.61083. Elife. 2020. PMID: 33084577 Free PMC article.
-
Dynamic readout of the Hh gradient in the Drosophila wing disc reveals pattern-specific tradeoffs between robustness and precision.Elife. 2024 Nov 7;13:e85755. doi: 10.7554/eLife.85755. Elife. 2024. PMID: 39508736 Free PMC article.
-
Formation and maintenance of morphogen gradients: an essential role for the endomembrane system in Drosophila melanogaster wing development.Fly (Austin). 2011 Jul-Sep;5(3):266-71. doi: 10.4161/fly.5.3.16542. Epub 2011 Jul 1. Fly (Austin). 2011. PMID: 21654212 Review.
Cited by
-
Synthetically programming natural cell-cell communication pathways for tissue engineering.Curr Opin Biomed Eng. 2024 Dec;32:100554. doi: 10.1016/j.cobme.2024.100554. Epub 2024 Aug 24. Curr Opin Biomed Eng. 2024. PMID: 40655075 Free PMC article.
-
Drosophila hedgehog signaling range and robustness depend on direct and sustained heparan sulfate interactions.Front Mol Biosci. 2023 Feb 22;10:1130064. doi: 10.3389/fmolb.2023.1130064. eCollection 2023. Front Mol Biosci. 2023. PMID: 36911531 Free PMC article.
-
Hedgehog on the Move: Glypican-Regulated Transport and Gradient Formation in Drosophila.Cells. 2024 Feb 27;13(5):418. doi: 10.3390/cells13050418. Cells. 2024. PMID: 38474382 Free PMC article. Review.
-
Molecularly Defined Glycocalyx Models Reveal AB5 Toxins Recognize Their Target Glycans Superselectively.JACS Au. 2025 May 20;5(6):2699-2712. doi: 10.1021/jacsau.5c00305. eCollection 2025 Jun 23. JACS Au. 2025. PMID: 40575306 Free PMC article.
-
Interactions of proteins with heparan sulfate.Essays Biochem. 2024 Dec 4;68(4):479-489. doi: 10.1042/EBC20230093. Essays Biochem. 2024. PMID: 38646914 Free PMC article. Review.
References
-
- Ingham PW, McMahon AP. Hedgehog signaling in animal development: paradigms and principles. Genes Dev. 2001;15:3059–3087. - PubMed
-
- Murone M, Rosenthal A, de Sauvage FJ. Sonic hedgehog signaling by the patched-smoothened receptor complex. Curr. Biol. 1999;9:76–84. - PubMed
-
- Hooper JE, Scott MP. Communicating with hedgehogs. Nat. Rev. Mol. Cell Biol. 2005;6:306–317. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
