

Role of Histone Tails and Single Strand DNA Breaks in Nucleosomal Arrest of RNA Polymerase
- PMID: 36768621
- PMCID: PMC9917218
- DOI: 10.3390/ijms24032295
Role of Histone Tails and Single Strand DNA Breaks in Nucleosomal Arrest of RNA Polymerase
Abstract
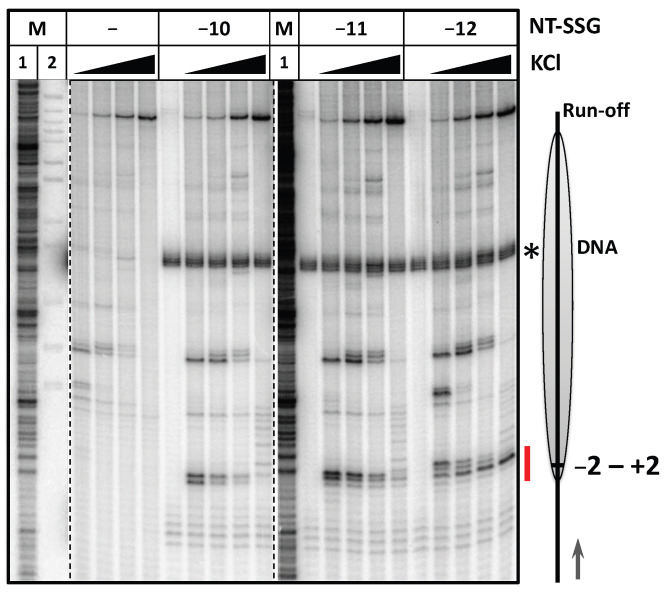
Transcription through nucleosomes by RNA polymerases (RNAP) is accompanied by formation of small intranucleosomal DNA loops (i-loops). The i-loops form more efficiently in the presence of single-strand breaks or gaps in a non-template DNA strand (NT-SSBs) and induce arrest of transcribing RNAP, thus allowing detection of NT-SSBs by the enzyme. Here we examined the role of histone tails and extranucleosomal NT-SSBs in i-loop formation and arrest of RNAP during transcription of promoter-proximal region of nucleosomal DNA. NT-SSBs present in linker DNA induce arrest of RNAP +1 to +15 bp in the nucleosome, suggesting formation of the i-loops; the arrest is more efficient in the presence of the histone tails. Consistently, DNA footprinting reveals formation of an i-loop after stalling RNAP at the position +2 and backtracking to position +1. The data suggest that histone tails and NT-SSBs present in linker DNA strongly facilitate formation of the i-loops during transcription through the promoter-proximal region of nucleosomal DNA.
Keywords: DNA damage; DNA loop; chromatin structure; nucleosome; single-strand DNA breaks; transcription-coupled DNA repair.
Conflict of interest statement
The authors declare no conflict of interest.
Figures






Similar articles
-
Structure of transcribed chromatin is a sensor of DNA damage.Sci Adv. 2015 Jul 3;1(6):e1500021. doi: 10.1126/sciadv.1500021. eCollection 2015 Jul. Sci Adv. 2015. PMID: 26601207 Free PMC article.
-
Transcription blockage by bulky end termini at single-strand breaks in the DNA template: differential effects of 5' and 3' adducts.Biochemistry. 2012 Nov 6;51(44):8964-70. doi: 10.1021/bi301240y. Epub 2012 Oct 24. Biochemistry. 2012. PMID: 23066636 Free PMC article.
-
Structure of an Intranucleosomal DNA Loop That Senses DNA Damage during Transcription.Cells. 2022 Aug 28;11(17):2678. doi: 10.3390/cells11172678. Cells. 2022. PMID: 36078089 Free PMC article.
-
Mechanism of transcription through a nucleosome by RNA polymerase II.Biochim Biophys Acta. 2013 Jan;1829(1):76-83. doi: 10.1016/j.bbagrm.2012.08.015. Epub 2012 Sep 6. Biochim Biophys Acta. 2013. PMID: 22982194 Free PMC article. Review.
-
[Molecular mechanisms of transcription through a nuclesome by RNA polymerase II].Mol Biol (Mosk). 2013 Sep-Oct;47(5):754-66. Mol Biol (Mosk). 2013. PMID: 25509347 Review. Russian.
Cited by
-
Structural Transition of the Nucleosome during Transcription Elongation.Cells. 2023 May 14;12(10):1388. doi: 10.3390/cells12101388. Cells. 2023. PMID: 37408222 Free PMC article. Review.
-
Histone tetrasome dynamics affects chromatin transcription.Nucleic Acids Res. 2025 Apr 22;53(8):gkaf356. doi: 10.1093/nar/gkaf356. Nucleic Acids Res. 2025. PMID: 40304183 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
