

Supt16 Haploinsufficiency Impairs PI3K/AKT/mTOR/Autophagy Pathway in Human Pluripotent Stem Cells Derived Neural Stem Cells
- PMID: 36769360
- PMCID: PMC9917370
- DOI: 10.3390/ijms24033035
Supt16 Haploinsufficiency Impairs PI3K/AKT/mTOR/Autophagy Pathway in Human Pluripotent Stem Cells Derived Neural Stem Cells
Abstract
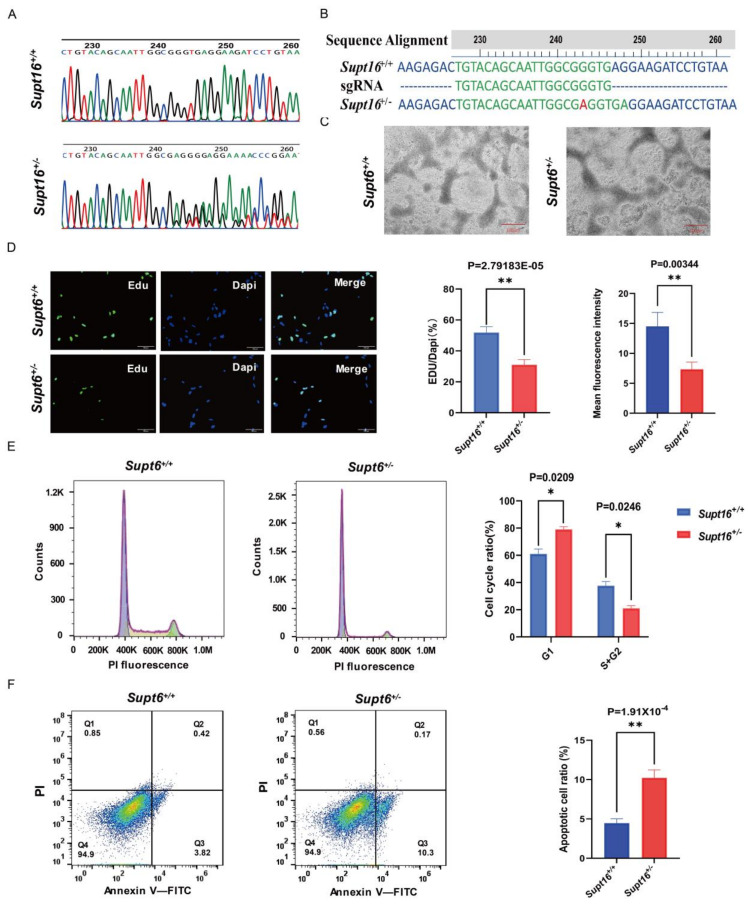
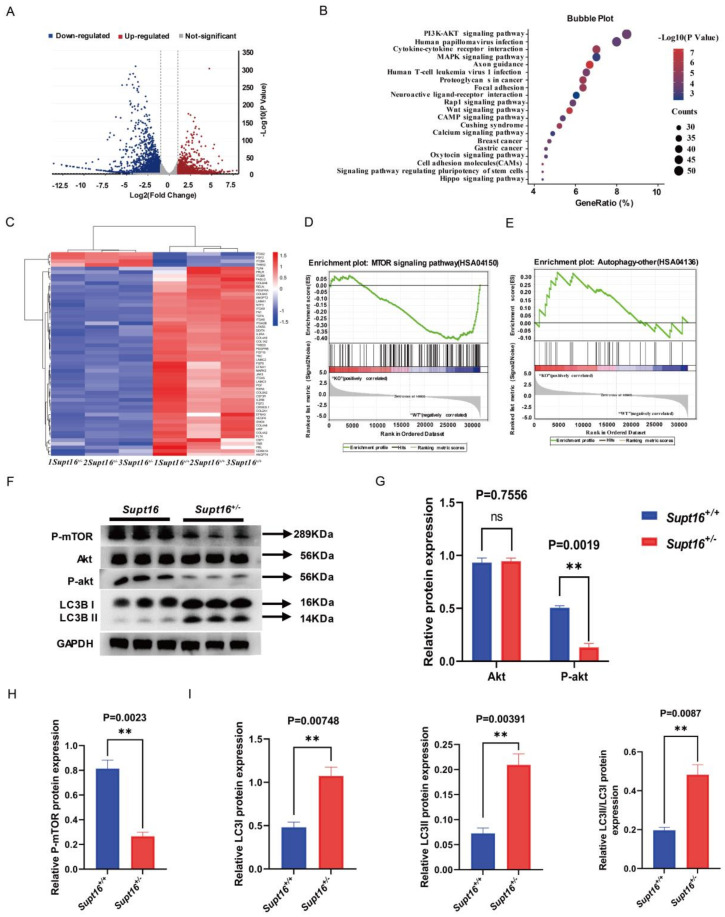
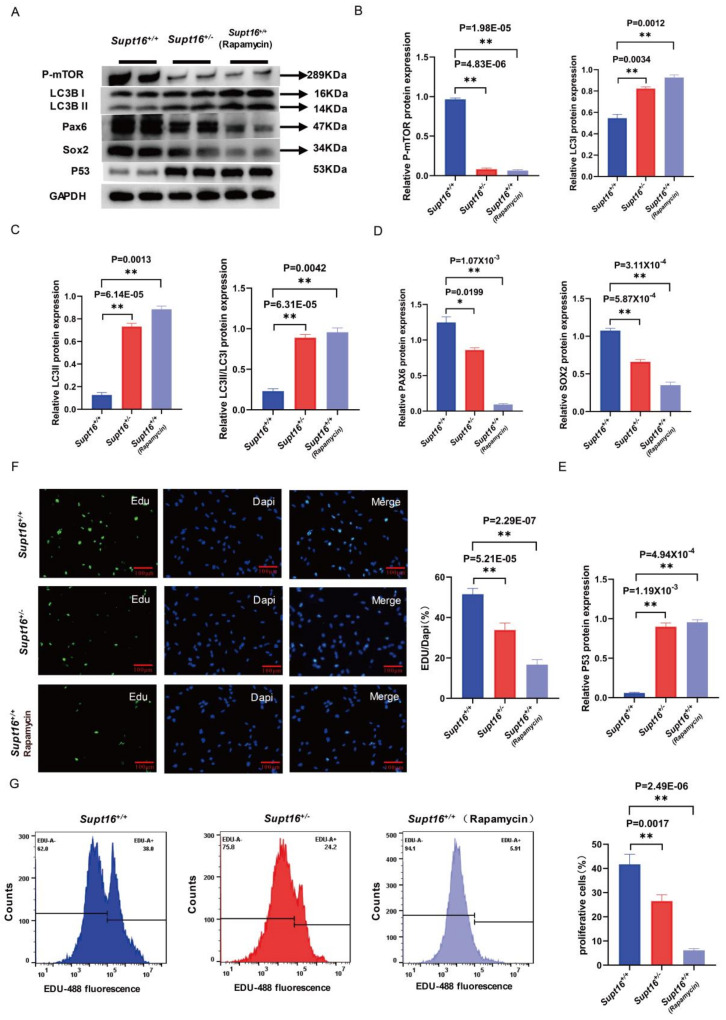
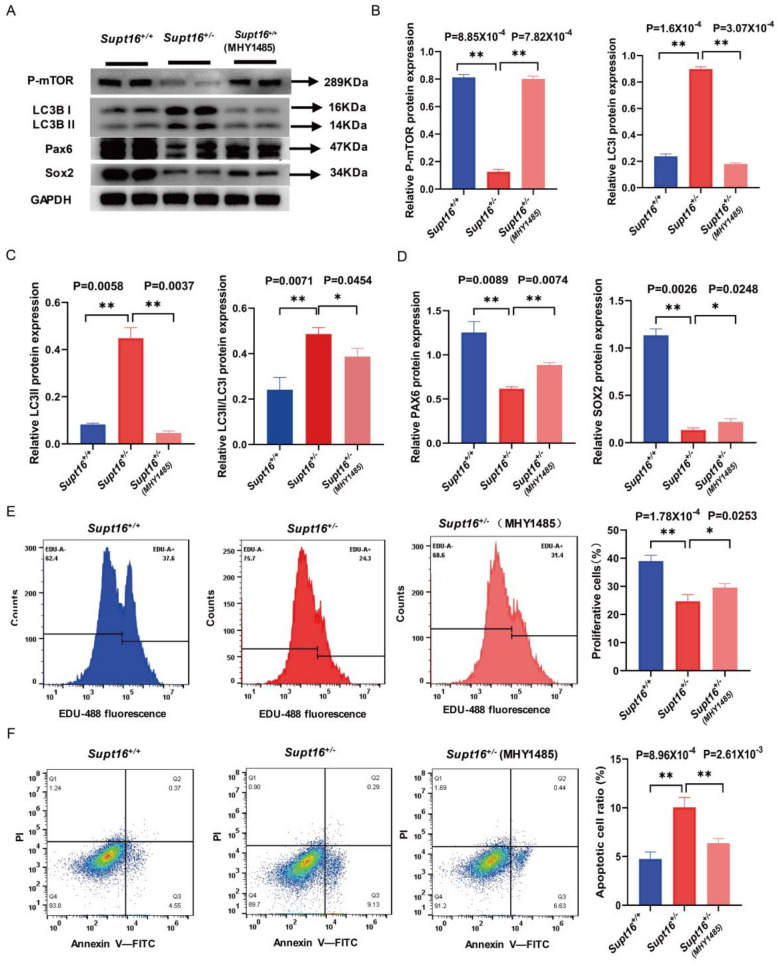
The maintenance of neural stem cells (NSCs) plays a critical role in neurodevelopment and has been implicated in neurodevelopmental disorders (NDDs). However, the underlying mechanisms linking defective human neural stem cell self-renewal to NDDs remain undetermined. Our previous study found that Supt16 haploinsufficiency causes cognitive and social behavior deficits by disrupting the stemness maintenance of NSCs in mice. However, its effects and underlying mechanisms have not been elucidated in human neural stem cells (hNSCs). Here, we generated Supt16+/- induced pluripotent stem cells (iPSCs) and induced them into hNSCs. The results revealed that Supt16 heterozygous hNSCs exhibit impaired proliferation, cell cycle arrest, and increased apoptosis. As the RNA-seq analysis showed, Supt16 haploinsufficiency inhibited the PI3K/AKT/mTOR pathway, leading to rising autophagy, and further resulted in the dysregulated expression of multiple proteins related to cell proliferation and apoptotic process. Furthermore, the suppression of Supt16 heterozygous hNSC self-renewal caused by autophagy activation could be rescued by MHY1485 treatment or reproduced in rapamycin-treated hNSCs. Thus, our results showed that Supt16 was essential for hNSC self-renewal and its haploinsufficiency led to cell cycle arrest, impaired cell proliferation, and increased apoptosis of hNSCs by regulating the PI3K/AKT/mTOR/autophagy pathway. These provided a new insight to understand the causality between the Supt16 heterozygous NSCs and NDDs in humans.
Keywords: Supt16; autophagy; human neural stem cells (hNSCs); neurodevelopmental disorders (NDDs); proliferation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
