

HDAC8 regulates protein kinase D phosphorylation in skeletal myoblasts in response to stress signaling
- PMID: 36773343
- PMCID: PMC9975084
- DOI: 10.1016/j.bbrc.2023.02.010
HDAC8 regulates protein kinase D phosphorylation in skeletal myoblasts in response to stress signaling
Abstract
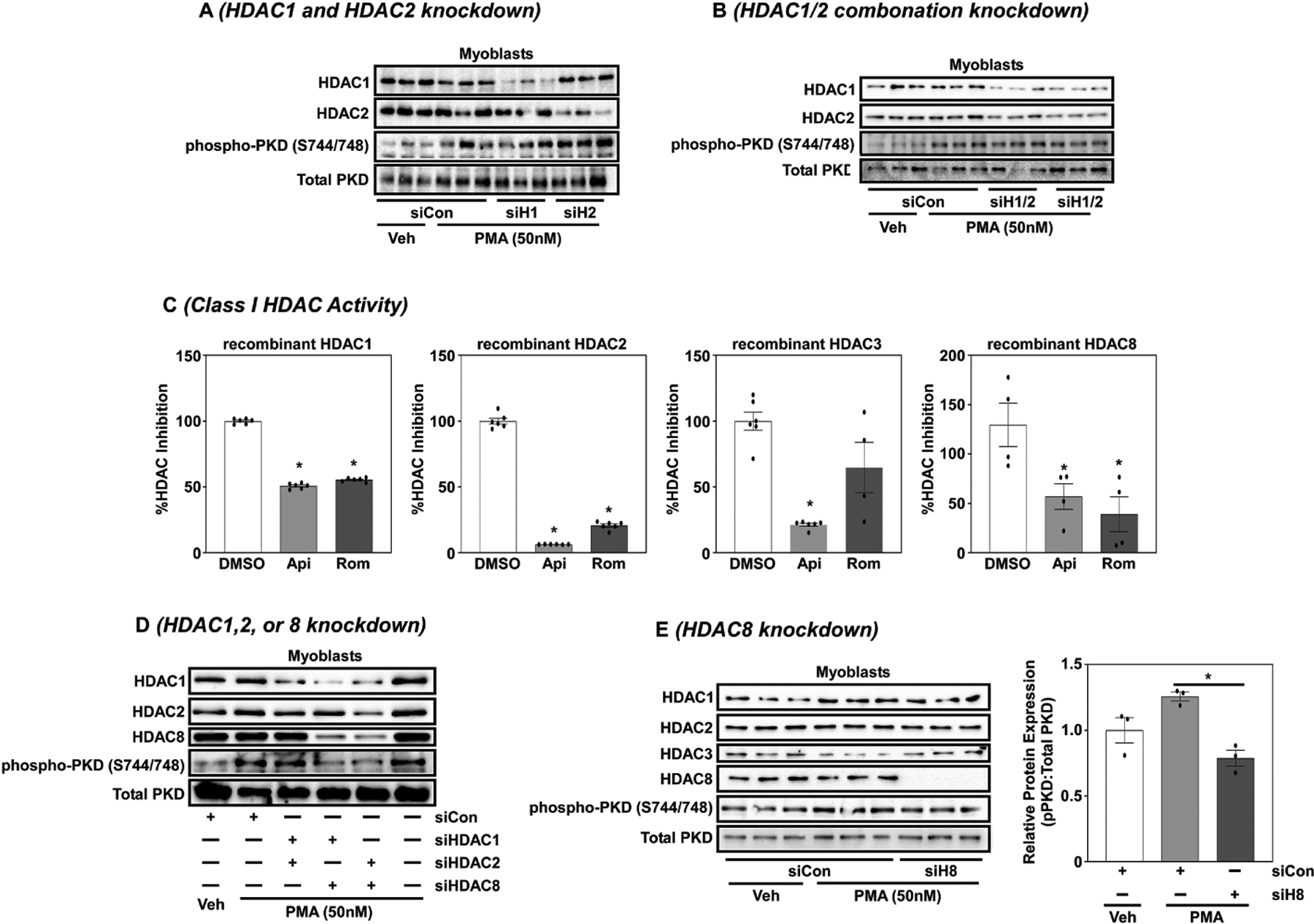
Skeletal muscle differentiation involves activation of quiescent satellite cells to proliferate, differentiate and fuse to form new myofibers; this requires coordination of myogenic transcription factors. Myogenic transcription is tightly regulated by various intracellular signaling pathways, which include members of the protein kinase D (PKD) family. PKD is a family of serine-threonine kinases that regulate gene expression, protein secretion, cell proliferation, differentiation and inflammation. PKD is a unique PKC family member that shares distant sequence homology to calcium-regulated kinases and plays an important role in muscle physiology. In this report, we show that class I histone deacetylase (HDAC) inhibition, and in particular HDAC8 inhibition, attenuated PKD phosphorylation in skeletal C2C12 myoblasts in response to phorbol ester, angiotensin II and dexamethasone signaling independent of changes in total PKD protein expression. As class I HDACs and PKD signaling are requisite for myocyte differentiation, these data suggest that HDAC8 functions as a potential feedback regulator of PKD phosphorylation to control myogenic gene expression.
Keywords: HDAC; HDAC inhibitors; Histone deacetylase; Myoblast; PKD; Protein kinase D.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of competing interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures


References
-
- Kleger A, Loebnitz C, Pusapati GV, Armacki M, ller MM, Tümpel S, Illing A, Hartmann D, Brunner C, Liebau S, Rudolph KL, Adler G, Seufferlein T, Protein Kinase D2 Is an Essential Regulator of Murine Myoblast Differentiation, PLoS One. 6 (2011) e14599. 10.1371/JOURNAL.PONE.0014599. - DOI - PMC - PubMed
-
- Czifra G, Tóth IB, Marincsák R, Juhász I, Kovács I, Ács P, Kovács L, Blumberg PM, Bíró T, Insulin-like growth factor-I-coupled mitogenic signaling in primary cultured human skeletal muscle cells and in C2C12 myoblasts. A central role of protein kinase Cδ, (2006). 10.1016/j.cellsig.2005.11.007. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
