

Acute and long-term exercise adaptation of adipose tissue and skeletal muscle in humans: a matched transcriptomics approach after 8-week training-intervention
- PMID: 36774413
- PMCID: PMC10113153
- DOI: 10.1038/s41366-023-01271-y
Acute and long-term exercise adaptation of adipose tissue and skeletal muscle in humans: a matched transcriptomics approach after 8-week training-intervention
Abstract
Background: Exercise exerts many health benefits by directly inducing molecular alterations in physically utilized skeletal muscle. Molecular adaptations of subcutaneous adipose tissue (SCAT) might also contribute to the prevention of metabolic diseases.
Aim: To characterize the response of human SCAT based on changes in transcripts and mitochondrial respiration to acute and repeated bouts of exercise in comparison to skeletal muscle.
Methods: Sedentary participants (27 ± 4 yrs) with overweight or obesity underwent 8-week supervised endurance exercise 3×1h/week at 80% VO2peak. Before, 60 min after the first and last exercise bout and 5 days post intervention, biopsies were taken for transcriptomic analyses and high-resolution respirometry (n = 14, 8 female/6 male).
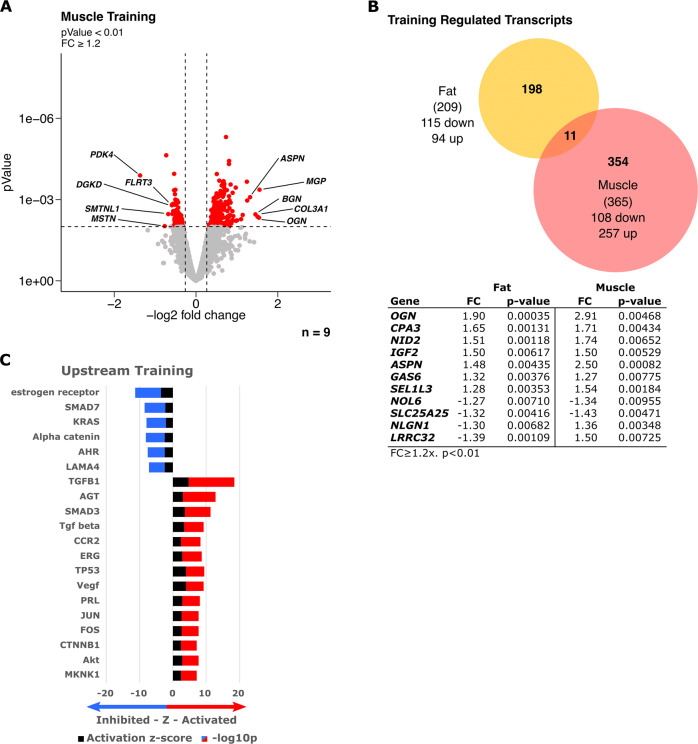
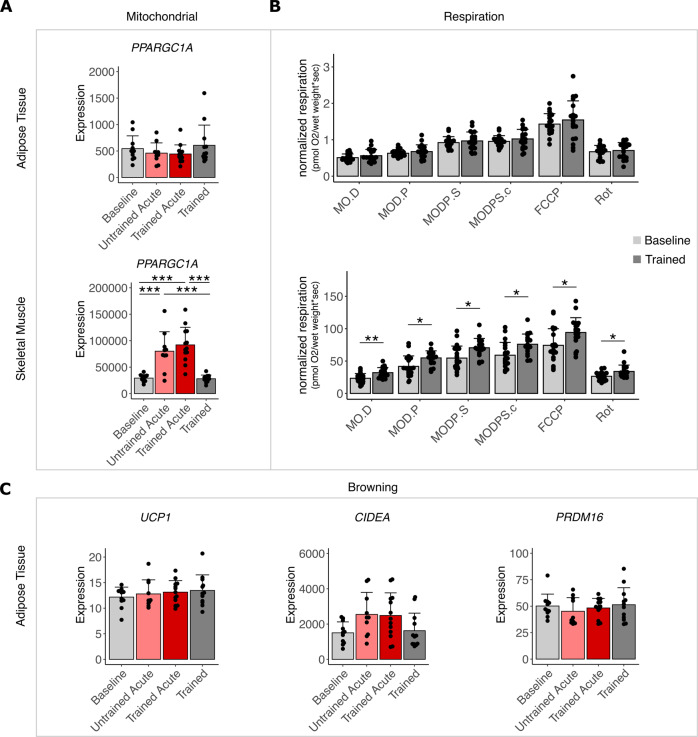
Results: In SCAT, we found 37 acutely regulated transcripts (FC > 1.2, FDR < 10%) after the first exercise bout compared to 394, respectively, in skeletal muscle. Regulation of only 5 transcripts overlapped between tissues highlighting their differential response. Upstream and enrichment analyses revealed reduced transcripts of lipid uptake, storage and lipogenesis directly after exercise in SCAT and point to β-adrenergic regulation as potential major driver. The data also suggest an exercise-induced modulation of the circadian clock in SCAT. Neither term was associated with transcriptomic changes in skeletal muscle. No evidence for beigeing/browning was found in SCAT along with unchanged respiration.
Conclusions: Adipose tissue responds completely distinct from adaptations of skeletal muscle to exercise. The acute and repeated reduction in transcripts of lipid storage and lipogenesis, interconnected with a modulated circadian rhythm, can counteract metabolic syndrome progression toward diabetes.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




Similar articles
-
Sex differences in resting skeletal muscle and the acute and long-term response to endurance exercise in individuals with overweight and obesity.Mol Metab. 2025 Aug;98:102185. doi: 10.1016/j.molmet.2025.102185. Epub 2025 Jun 16. Mol Metab. 2025. PMID: 40516819 Free PMC article.
-
Exercise rehabilitation following intensive care unit discharge for recovery from critical illness.Cochrane Database Syst Rev. 2015 Jun 22;2015(6):CD008632. doi: 10.1002/14651858.CD008632.pub2. Cochrane Database Syst Rev. 2015. PMID: 26098746 Free PMC article.
-
Physical exercise training interventions for children and young adults during and after treatment for childhood cancer.Cochrane Database Syst Rev. 2016 Mar 31;3(3):CD008796. doi: 10.1002/14651858.CD008796.pub3. Cochrane Database Syst Rev. 2016. PMID: 27030386 Free PMC article.
-
Physical exercise training interventions for children and young adults during and after treatment for childhood cancer.Cochrane Database Syst Rev. 2013 Apr 30;(4):CD008796. doi: 10.1002/14651858.CD008796.pub2. Cochrane Database Syst Rev. 2013. Update in: Cochrane Database Syst Rev. 2016 Mar 31;3:CD008796. doi: 10.1002/14651858.CD008796.pub3. PMID: 23633361 Updated.
-
Comparison of Two Modern Survival Prediction Tools, SORG-MLA and METSSS, in Patients With Symptomatic Long-bone Metastases Who Underwent Local Treatment With Surgery Followed by Radiotherapy and With Radiotherapy Alone.Clin Orthop Relat Res. 2024 Dec 1;482(12):2193-2208. doi: 10.1097/CORR.0000000000003185. Epub 2024 Jul 23. Clin Orthop Relat Res. 2024. PMID: 39051924
Cited by
-
Accelerometer-derived "weekend warrior" physical activity, sedentary behavior, and risk of dementia.Alzheimers Res Ther. 2025 Mar 22;17(1):67. doi: 10.1186/s13195-024-01657-8. Alzheimers Res Ther. 2025. PMID: 40121491 Free PMC article.
-
Mitochondrial Function in Healthy Human White Adipose Tissue: A Narrative Review.Nutrients. 2023 Oct 19;15(20):4430. doi: 10.3390/nu15204430. Nutrients. 2023. PMID: 37892505 Free PMC article. Review.
-
Skeletal Muscle Gene Expression Signatures of Obese High and Low Responders to Endurance Exercise Training.J Clin Endocrinol Metab. 2024 Apr 19;109(5):1318-1327. doi: 10.1210/clinem/dgad677. J Clin Endocrinol Metab. 2024. PMID: 37988600 Free PMC article.
-
Skeletal muscle BMAL1 is necessary for transcriptional adaptation of local and peripheral tissues in response to endurance exercise training.Mol Metab. 2024 Aug;86:101980. doi: 10.1016/j.molmet.2024.101980. Epub 2024 Jun 29. Mol Metab. 2024. PMID: 38950777 Free PMC article.
-
The role of adipose tissue dysfunction in hepatic insulin resistance and T2D.J Endocrinol. 2024 Aug 2;262(3):e240115. doi: 10.1530/JOE-24-0115. Print 2024 Sep 1. J Endocrinol. 2024. PMID: 38967989 Free PMC article. Review.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
