

Principles of target DNA cleavage and the role of Mg2+ in the catalysis of CRISPR-Cas9
- PMID: 36778082
- PMCID: PMC9909973
- DOI: 10.1038/s41929-022-00848-6
Principles of target DNA cleavage and the role of Mg2+ in the catalysis of CRISPR-Cas9
Abstract
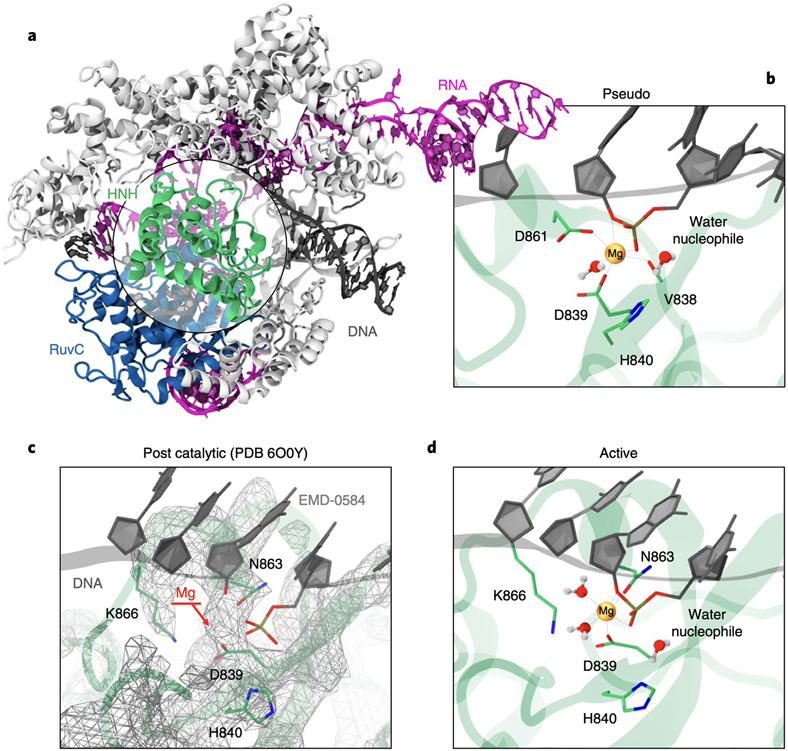
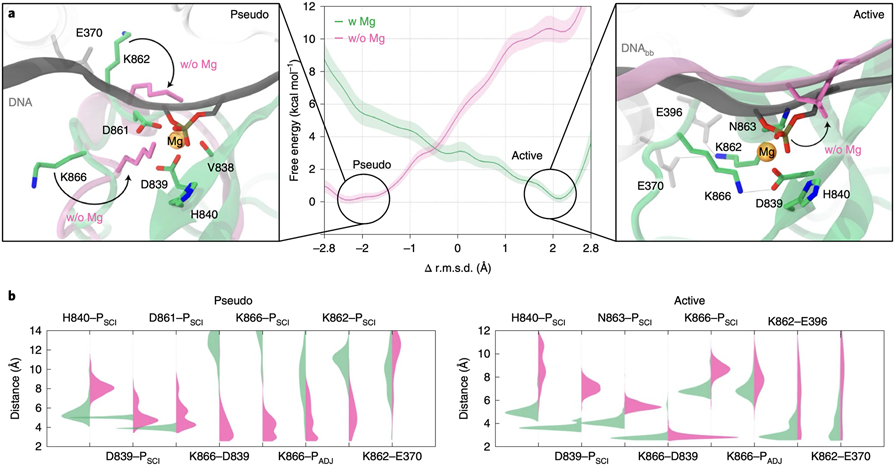
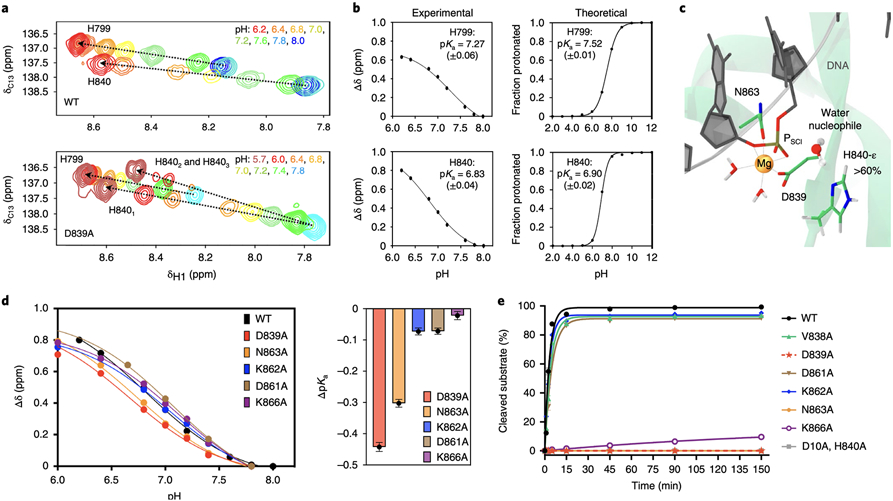
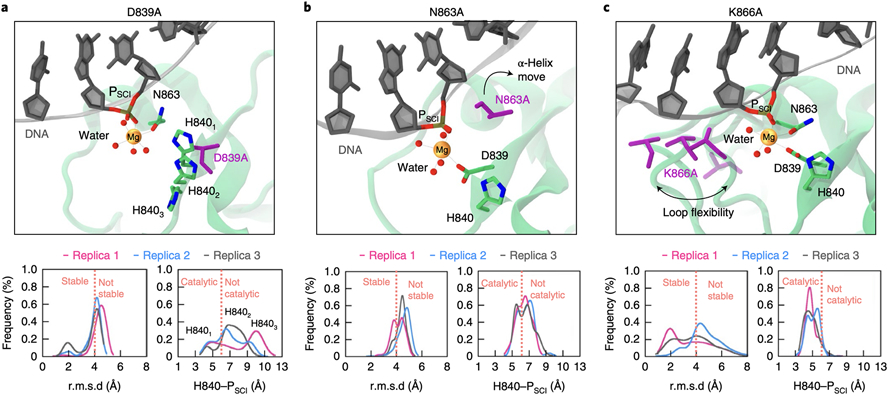
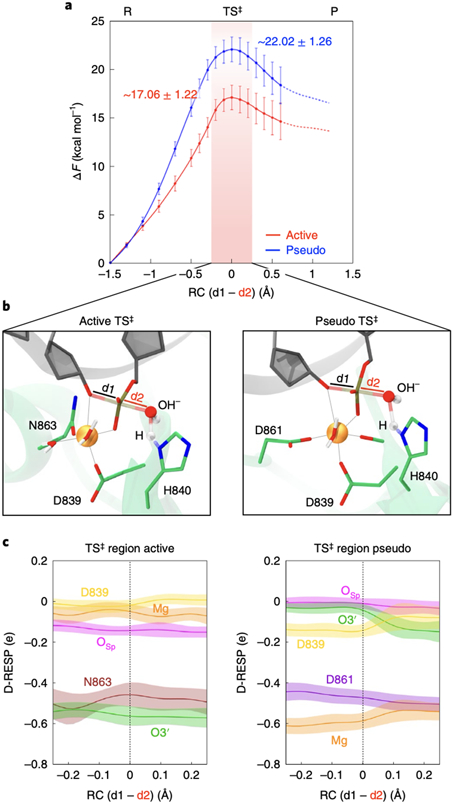
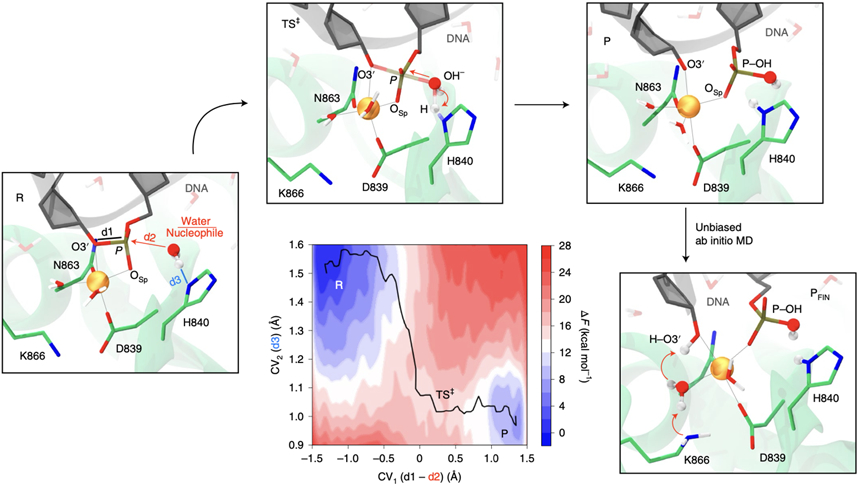
At the core of the CRISPR-Cas9 genome-editing technology, the endonuclease Cas9 introduces site-specific breaks in DNA. However, precise mechanistic information to ameliorating Cas9 function is still missing. Here, multi-microsecond molecular dynamics, free-energy and multiscale simulations are combined with solution NMR and DNA cleavage experiments to resolve the catalytic mechanism of target DNA cleavage. We show that the conformation of an active HNH nuclease is tightly dependent on the catalytic Mg2+, unveiling its cardinal structural role. This activated Mg2+-bound HNH is consistently described through molecular simulations, solution NMR and DNA cleavage assays, revealing also that the protonation state of the catalytic H840 is strongly affected by active site mutations. Finally, ab-initio QM(DFT)/MM simulations and metadynamics establish the catalytic mechanism, showing that the catalysis is activated by H840 and completed by K866, rationalising DNA cleavage experiments. This information is critical to enhance the enzymatic function of CRISPR-Cas9 toward improved genome-editing.
Figures
Similar articles
-
Structural and functional insights into the bona fide catalytic state of Streptococcus pyogenes Cas9 HNH nuclease domain.Elife. 2019 Jul 30;8:e46500. doi: 10.7554/eLife.46500. Elife. 2019. PMID: 31361218 Free PMC article.
-
Catalytic Mechanism of Non-Target DNA Cleavage in CRISPR-Cas9 Revealed by Ab Initio Molecular Dynamics.ACS Catal. 2020 Nov 20;10(22):13596-13605. doi: 10.1021/acscatal.0c03566. Epub 2020 Nov 10. ACS Catal. 2020. PMID: 33520346 Free PMC article.
-
Exploring CRISPR-Cas9 HNH-Domain-Catalyzed DNA Cleavage Using Accelerated Quantum Mechanical Molecular Mechanical Free Energy Simulation.Biochemistry. 2025 Jan 7;64(1):289-299. doi: 10.1021/acs.biochem.4c00651. Epub 2024 Dec 16. Biochemistry. 2025. PMID: 39680038
-
Dynamics and mechanisms of CRISPR-Cas9 through the lens of computational methods.Curr Opin Struct Biol. 2022 Aug;75:102400. doi: 10.1016/j.sbi.2022.102400. Epub 2022 Jun 8. Curr Opin Struct Biol. 2022. PMID: 35689914 Free PMC article. Review.
-
Insights into the Mechanism of CRISPR/Cas9-Based Genome Editing from Molecular Dynamics Simulations.ACS Omega. 2022 Dec 30;8(2):1817-1837. doi: 10.1021/acsomega.2c05583. eCollection 2023 Jan 17. ACS Omega. 2022. PMID: 36687047 Free PMC article. Review.
Cited by
-
Observing one-divalent-metal-ion dependent and histidine-promoted His-Me family I-PpoI nuclease catalysis in crystallo.bioRxiv [Preprint]. 2024 Jul 11:2024.05.02.592236. doi: 10.1101/2024.05.02.592236. bioRxiv. 2024. Update in: Elife. 2024 Aug 14;13:RP99960. doi: 10.7554/eLife.99960. PMID: 38746211 Free PMC article. Updated. Preprint.
-
Increasing the Activity of the High-Fidelity SpyCas9 Form in Yeast by Directed Mutagenesis of the PAM-Interacting Domain.Int J Mol Sci. 2023 Dec 28;25(1):444. doi: 10.3390/ijms25010444. Int J Mol Sci. 2023. PMID: 38203615 Free PMC article.
-
Structural insights into Type II-D Cas9 and its robust cleavage activity.Nat Commun. 2025 Aug 11;16(1):7396. doi: 10.1038/s41467-025-62128-8. Nat Commun. 2025. PMID: 40790018 Free PMC article.
-
Disruption of electrostatic contacts in the HNH nuclease from a thermophilic Cas9 rewires allosteric motions and enhances high-temperature DNA cleavage.J Chem Phys. 2022 Dec 14;157(22):225103. doi: 10.1063/5.0128815. J Chem Phys. 2022. PMID: 36546784 Free PMC article.
-
Cloning, heterologous expression, and molecular characterization of a highly active and stable non-specific endonuclease from Pseudomonas fluorescens.Arch Microbiol. 2024 Feb 27;206(3):125. doi: 10.1007/s00203-024-03867-y. Arch Microbiol. 2024. PMID: 38411841
References
-
- Doudna JA & Charpentier E Genome editing. The new frontier of genome engineering with CRISPR-Cas9. Science 346, 1258096 (2014). - PubMed