

This is a preprint.
Single-cell RNA sequencing reveals dysregulated fibroblast subclusters in prurigo nodularis
- PMID: 36778229
- PMCID: PMC9915465
- DOI: 10.1101/2023.01.29.526050
Single-cell RNA sequencing reveals dysregulated fibroblast subclusters in prurigo nodularis
Update in
-
Single-Cell RNA Sequencing Reveals Dysregulated POSTN+WNT5A+ Fibroblast Subclusters in Prurigo Nodularis.J Invest Dermatol. 2024 Jul;144(7):1568-1578.e5. doi: 10.1016/j.jid.2023.12.021. Epub 2024 Jan 20. J Invest Dermatol. 2024. PMID: 38246584
Abstract
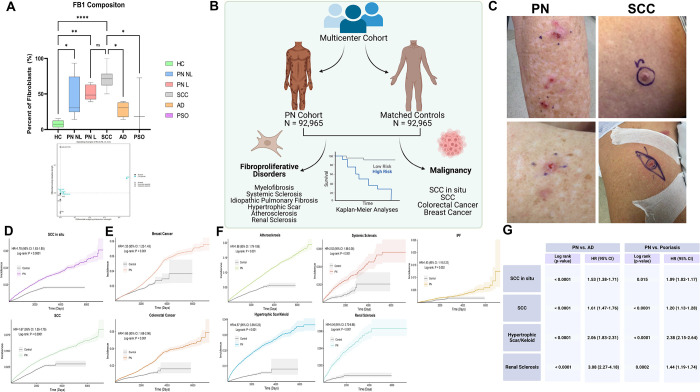
Prurigo nodularis (PN) is an intensely pruritic, chronic inflammatory skin disease that disproportionately affects black patients. However, the pathogenesis of PN is poorly understood. We performed single-cell transcriptomic profiling, ligand receptor analysis and cell trajectory analysis of 28,695 lesional and non-lesional PN skin cells to uncover disease-identifying cell compositions and genetic characteristics. We uncovered a dysregulated role for fibroblasts (FBs) and myofibroblasts as a key pathogenic element in PN, which were significantly increased in PN lesional skin. We defined seven unique subclusters of FBs in PN skin and observed a shift of PN lesional FBs towards a cancer-associated fibroblast (CAF)-like phenotype, with WNT5A+ CAFs increased in the skin of PN patients and similarly so in squamous cell carcinoma (SCC). A multicenter PN cohort study subsequently revealed an increased risk of SCC as well as additional CAF-associated malignancies in PN patients, including breast and colorectal cancers. Systemic fibroproliferative diseases were also upregulated in PN patients, including renal sclerosis and idiopathic pulmonary fibrosis. Ligand receptor analyses demonstrated increased FB1-derived WNT5A and periostin interactions with neuronal receptors MCAM and ITGAV, suggesting a fibroblast-neuronal axis in PN. Type I IFN responses in immune cells and increased angiogenesis/permeability in endothelial cells were also observed. As compared to atopic dermatitis (AD) and psoriasis (PSO) patients, increased mesenchymal dysregulation is unique to PN with an intermediate Th2/Th17 phenotype between atopic dermatitis and psoriasis. These findings identify a pathogenic role for CAFs in PN, including a novel targetable WNT5A+ fibroblast subpopulation and CAF-associated malignancies in PN patients.
Conflict of interest statement
Competing interests: Dr. Kwatra is an advisory board member/consultant for Abbvie, Aslan Pharmaceuticals, Arcutis Biotherapeutics, Celldex Therapeutics, Castle Biosciences, Galderma, Genzada Pharmaceuticals, Incyte Corporation, Johnson & Johnson, Leo Pharma, Novartis Pharmaceuticals Corporation, Pfizer, Regeneron Pharmaceuticals, and Sanofi and has served as an investigator for Galderma, Incyte, Pfizer, and Sanofi.
Figures







References
-
- Kwatra S.G. Breaking the Itch-Scratch Cycle in Prurigo Nodularis. N Engl J Med 382, 757–758 (2020). - PubMed
-
- Whang K.A., et al. Health-related quality of life and economic burden of prurigo nodularis. Journal of the American Academy of Dermatology 86, 573–580 (2022). - PubMed
-
- Huang A.H., Canner J.K., Khanna R., Kang S. & Kwatra S.G. Real-world prevalence of prurigo nodularis and burden of associated diseases. Journal of Investigative Dermatology 140, 480–483. e484 (2020). - PubMed
-
- Sutaria N., et al. Racial differences in dysregulation of the renin-angiotensin-aldosterone system in patients with prurigo nodularis. Journal of Dermatological Science 105, 130–136 (2022). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials