

This is a preprint.
A de novo missense variant in EZH1 associated with developmental delay exhibits functional deficits in Drosophila melanogaster
- PMID: 36778246
- PMCID: PMC9915809
- DOI: 10.1101/2023.01.31.23285113
A de novo missense variant in EZH1 associated with developmental delay exhibits functional deficits in Drosophila melanogaster
Update in
-
A de novo missense variant in EZH1 associated with developmental delay exhibits functional deficits in Drosophila melanogaster.Genetics. 2023 Aug 9;224(4):iyad110. doi: 10.1093/genetics/iyad110. Genetics. 2023. PMID: 37314226 Free PMC article.
Abstract
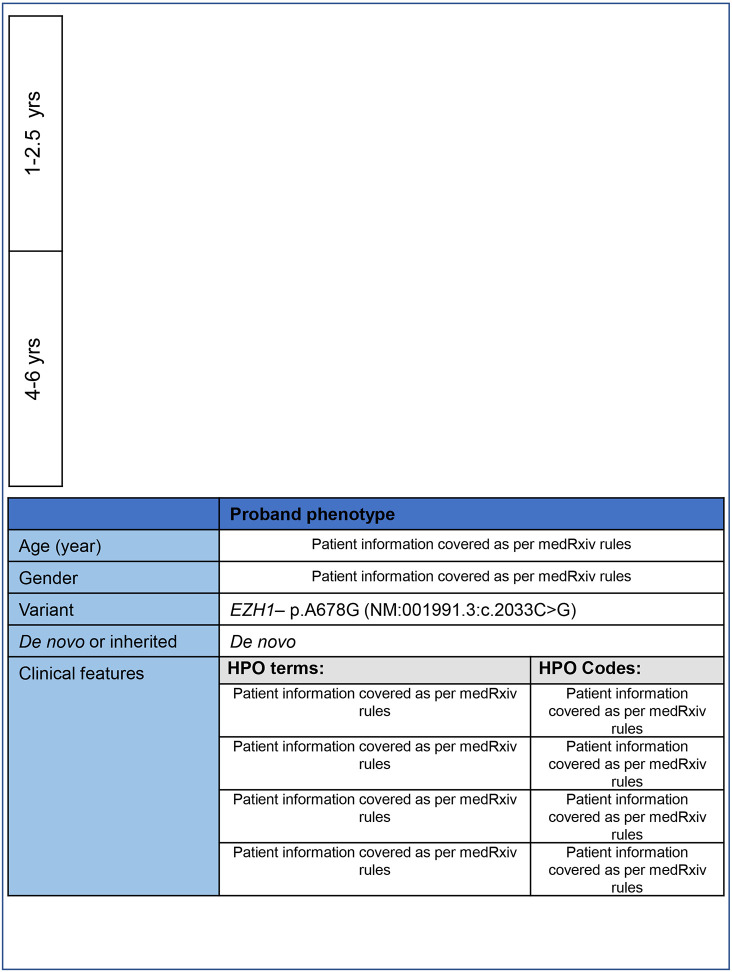
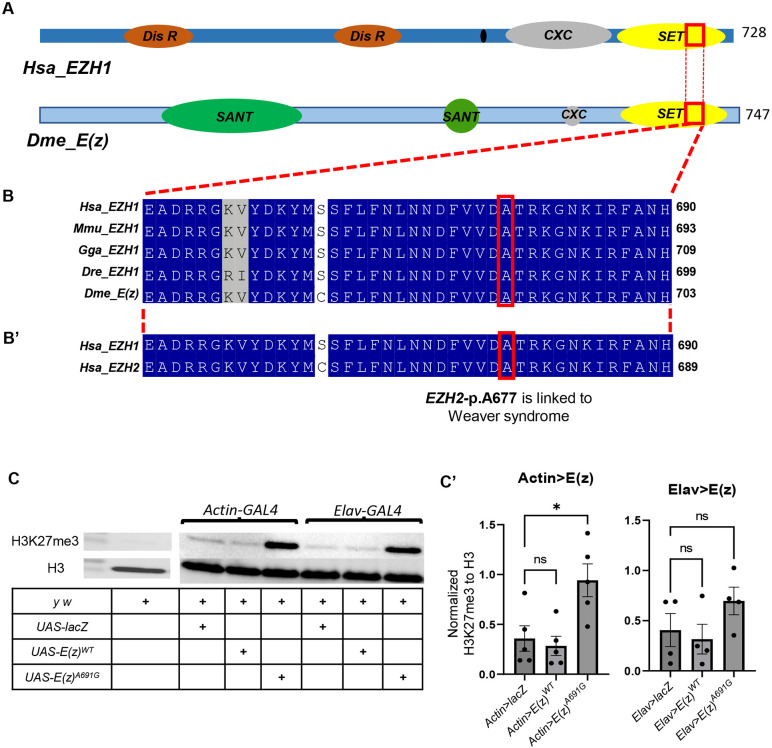
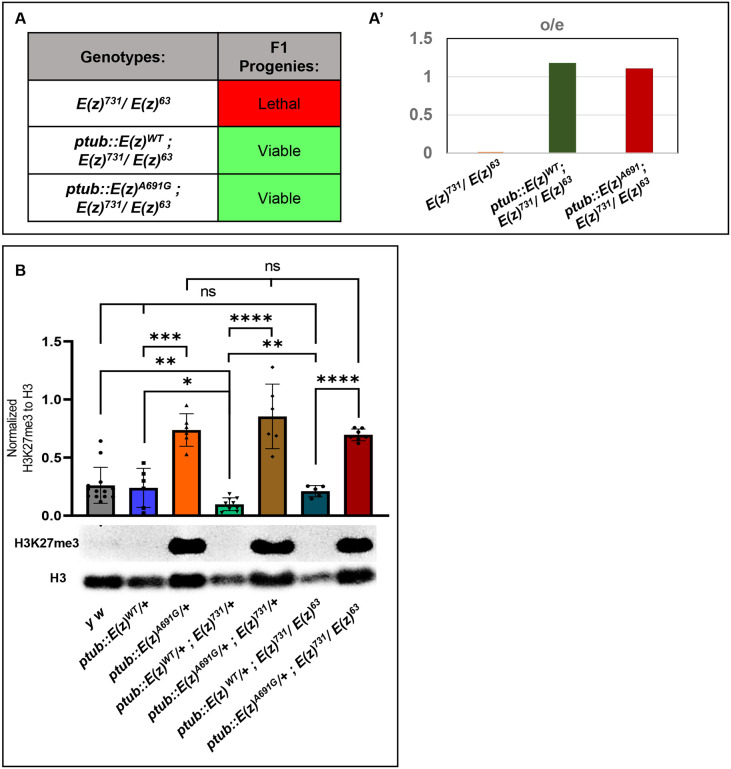
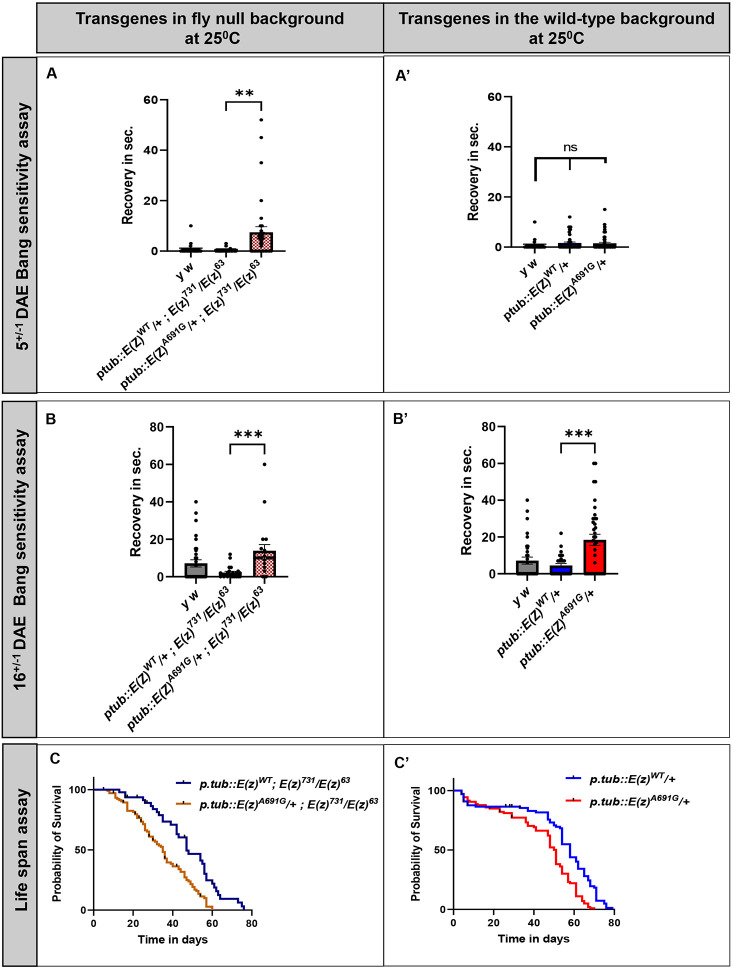
EZH1 ( Enhancer of Zeste, homolog 1) , a Polycomb Repressive Complex-2 (PRC2) component, is involved in a myriad of cellular processes through modifying histone 3 lysine27 (H3K27) residues. EZH1 represses transcription of downstream target genes through H3K27 trimethylation (H3K27me3). Genetic mutations in histone modifiers have been associated with developmental disorders, while EZH1 has not yet been linked to any human disease. However, the paralog EZH2 is associated with Weaver syndrome. Here we report a previously undiagnosed individual with a novel neurodevelopmental phenotype identified to have a de novo variant in EZH1 , p.Ala678Gly, through exome sequencing. The individual presented in infancy with neurodevelopmental delay and hypotonia and was later noted to have proximal muscle weakness. The variant, p.A678G, is in the SET domain, known for its methyltransferase activity, and was the best candidate variant found in the exome. Human EZH1 / 2 are homologous to fly Enhancer of zeste E(z) , an essential gene in flies, and the residue (A678 in humans, A691 in Drosophila ) is conserved. To further study this variant, we obtained Drosophila null alleles and generated transgenic flies expressing wild-type (E(z) WT ) and the variant (E(z) A691G ) . The E(z) A691G variant led to hyper H3K27me3 while the E(z) WT did not, suggesting this is as a gain-of-function allele. When expressed under the tubulin promotor in vivo the variant rescued null-lethality similar to wild-type but the E(z) A691G flies exhibit bang sensitivity and shortened lifespan. In conclusion, here we present a novel EZH1 de novo variant associated with a neurodevelopmental disorder. Furthermore, we found that this variant has a functional impact in Drosophila . Biochemically this allele leads to increased H3K27me3 suggesting gain-of-function, but when expressed in adult flies the E(z) A691G has some characteristics of partial loss-of-function which may suggest it is a more complex allele in vivo .
Figures
Similar articles
-
A de novo missense variant in EZH1 associated with developmental delay exhibits functional deficits in Drosophila melanogaster.Genetics. 2023 Aug 9;224(4):iyad110. doi: 10.1093/genetics/iyad110. Genetics. 2023. PMID: 37314226 Free PMC article.
-
Characterization of EZH1, a human homolog of Drosophila Enhancer of zeste near BRCA1.Genomics. 1996 Oct 15;37(2):161-71. doi: 10.1006/geno.1996.0537. Genomics. 1996. PMID: 8921387
-
Ezh1 and Ezh2 differentially regulate PSD-95 gene transcription in developing hippocampal neurons.Mol Cell Neurosci. 2013 Nov;57:130-43. doi: 10.1016/j.mcn.2013.07.012. Epub 2013 Aug 8. Mol Cell Neurosci. 2013. PMID: 23932971
-
Fibrosis-the tale of H3K27 histone methyltransferases and demethylases.Front Cell Dev Biol. 2023 Jul 5;11:1193344. doi: 10.3389/fcell.2023.1193344. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 37476157 Free PMC article. Review.
-
EZH1/2 as targets for cancer therapy.Cancer Gene Ther. 2023 Feb;30(2):221-235. doi: 10.1038/s41417-022-00555-1. Epub 2022 Nov 11. Cancer Gene Ther. 2023. PMID: 36369341 Review.
References
-
- Baldridge D, Undiagnosed Diseases Network, Wangler MF, Bowman AN, Yamamoto S, Schedl T, Pak SC, Postlethwait JH, Shin J, Solnica-Krezel L, et al. 2021. Model organisms contribute to diagnosis and discovery in the undiagnosed diseases network: current state and a future vision. Orphanet J Rare Dis 16. 10.1186/s13023-021-01839-9. - DOI - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources