

This is a preprint.
Replicative Acinetobacter baumannii strains interfere with phagosomal maturation by modulating the vacuolar pH
- PMID: 36778331
- PMCID: PMC9915592
- DOI: 10.1101/2023.02.02.526753
Replicative Acinetobacter baumannii strains interfere with phagosomal maturation by modulating the vacuolar pH
Update in
-
Replicative Acinetobacter baumannii strains interfere with phagosomal maturation by modulating the vacuolar pH.PLoS Pathog. 2023 Jun 9;19(6):e1011173. doi: 10.1371/journal.ppat.1011173. eCollection 2023 Jun. PLoS Pathog. 2023. PMID: 37294840 Free PMC article.
Abstract
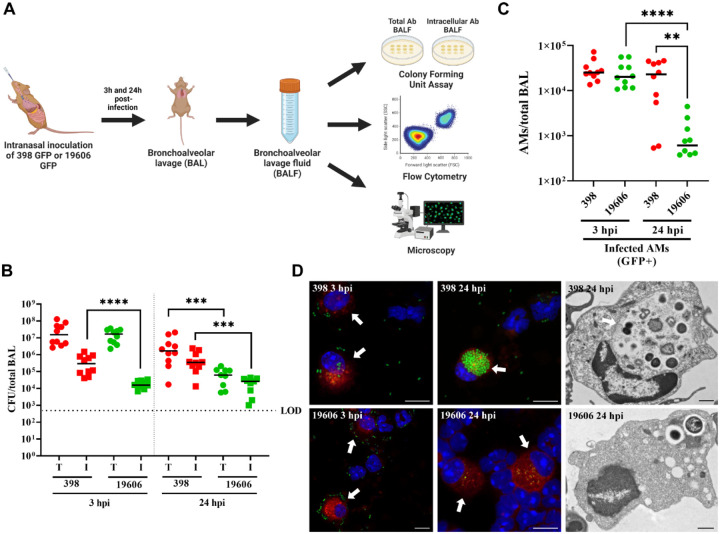
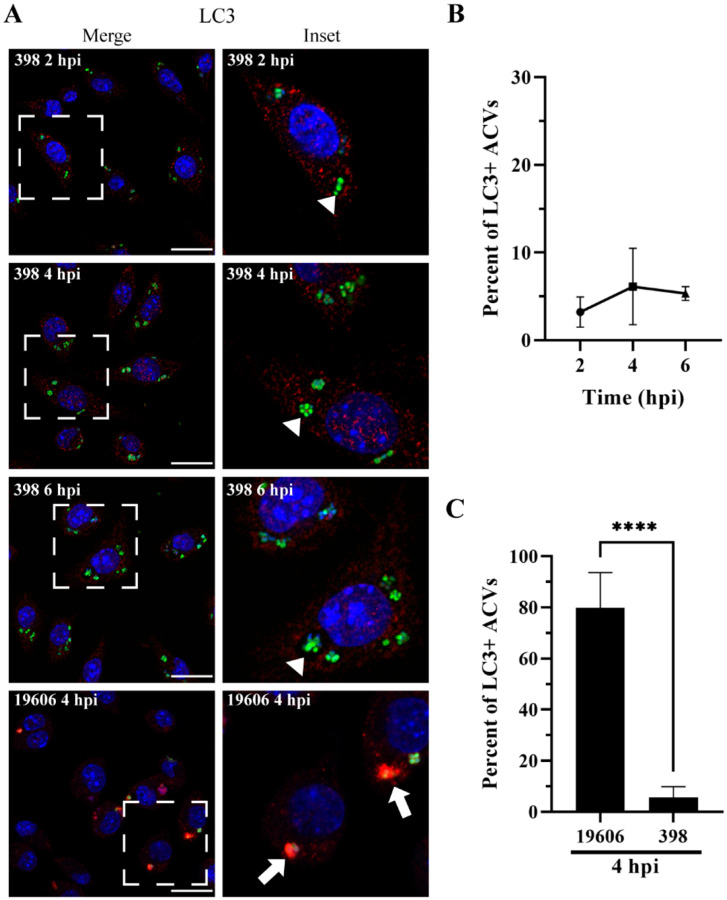
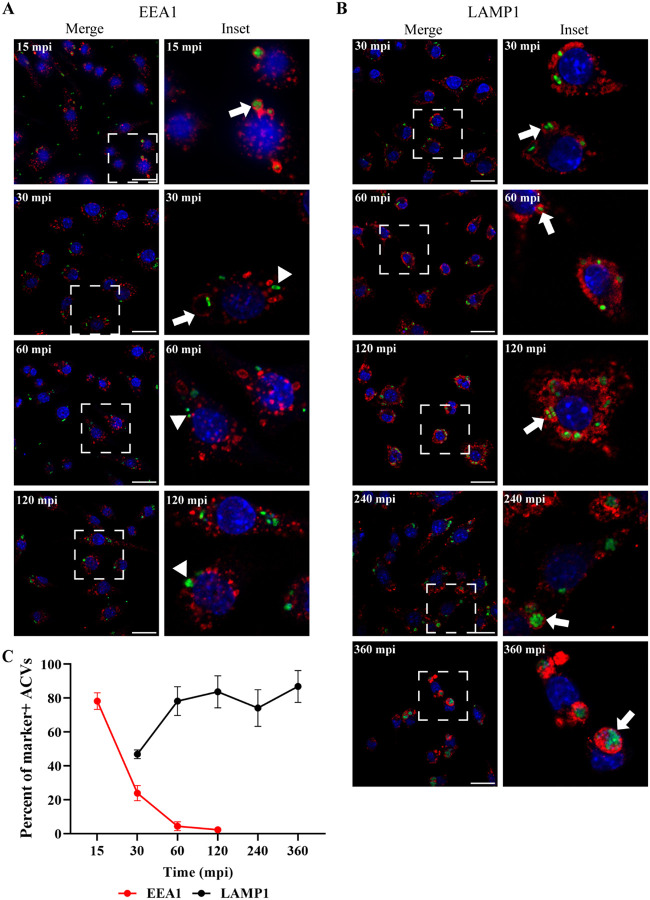
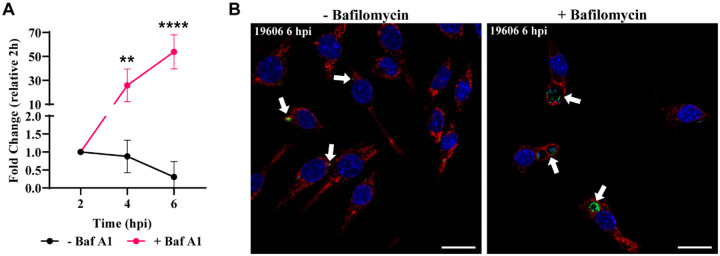
Bacterial pneumonia is a common infection of the lower respiratory tract that can afflict patients of all ages. Multidrug-resistant strains of Acinetobacter baumannii are increasingly responsible for causing nosocomial pneumonias, thus posing an urgent threat. Alveolar macrophages play a critical role in overcoming respiratory infections caused by this pathogen. Recently, we and others have shown that new clinical isolates of A. baumannii , but not the common lab strain ATCC 19606 (19606), can persist and replicate in macrophages within spacious vacuoles that we called A cinetobacter C ontaining V acuoles (ACV). In this work, we demonstrate that the modern A. baumannii clinical isolate 398, but not the lab strain 19606, can infect alveolar macrophages and produce ACVs in vivo in a murine pneumonia model. Both strains initially interact with the alveolar macrophage endocytic pathway, as indicated by EEA1 and LAMP1 markers; however, the fate of these strains diverges at a later stage. While 19606 is eliminated in an autophagy pathway, 398 replicates in ACVs and are not degraded. We show that 398 reverts the natural acidification of the phagosome by secreting large amounts of ammonia, a by-product of amino acid catabolism. We propose that this ability to survive within macrophages may be critical for the persistence of clinical A. baumannii isolates in the lung during a respiratory infection.
Conflict of interest statement
Competing Interest Statement: The authors declare no conflict of interest.
Figures




References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous