

This is a preprint.
Unstable EBV latency drives inflammation in multiple sclerosis patient derived spontaneous B cells
- PMID: 36778367
- PMCID: PMC9915775
- DOI: 10.21203/rs.3.rs-2398872/v1
Unstable EBV latency drives inflammation in multiple sclerosis patient derived spontaneous B cells
Update in
-
Multiple sclerosis patient-derived spontaneous B cells have distinct EBV and host gene expression profiles in active disease.Nat Microbiol. 2024 Jun;9(6):1540-1554. doi: 10.1038/s41564-024-01699-6. Epub 2024 May 28. Nat Microbiol. 2024. PMID: 38806670 Free PMC article.
Abstract
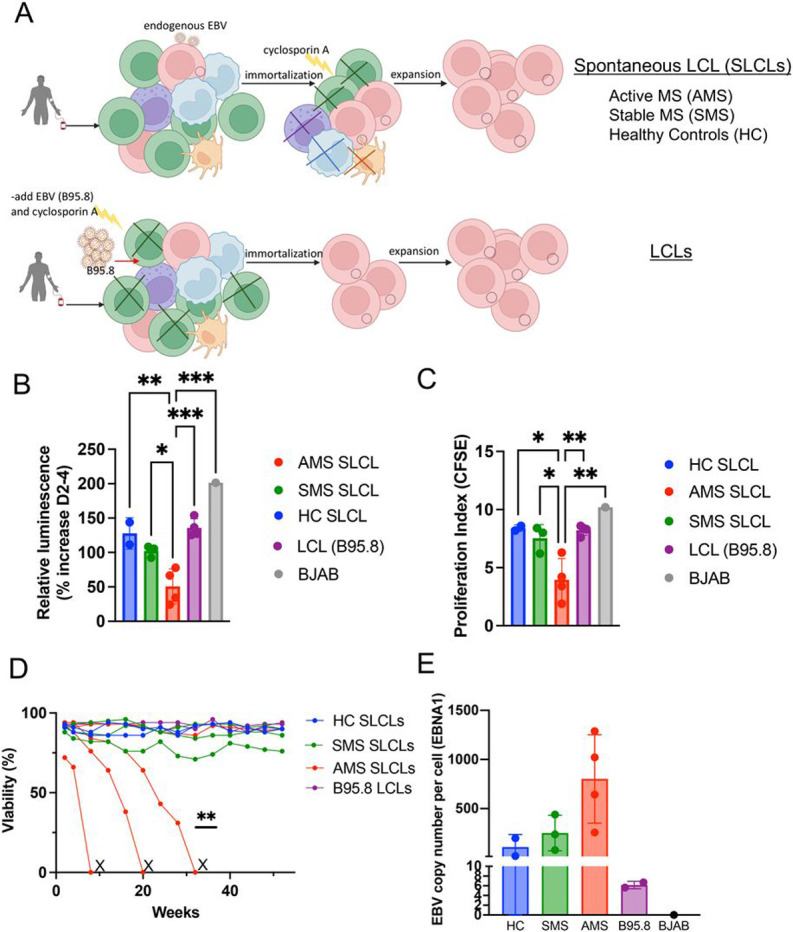
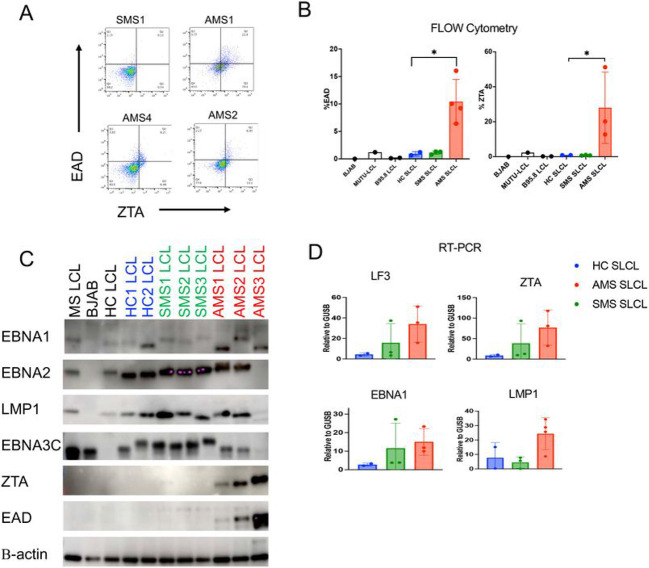
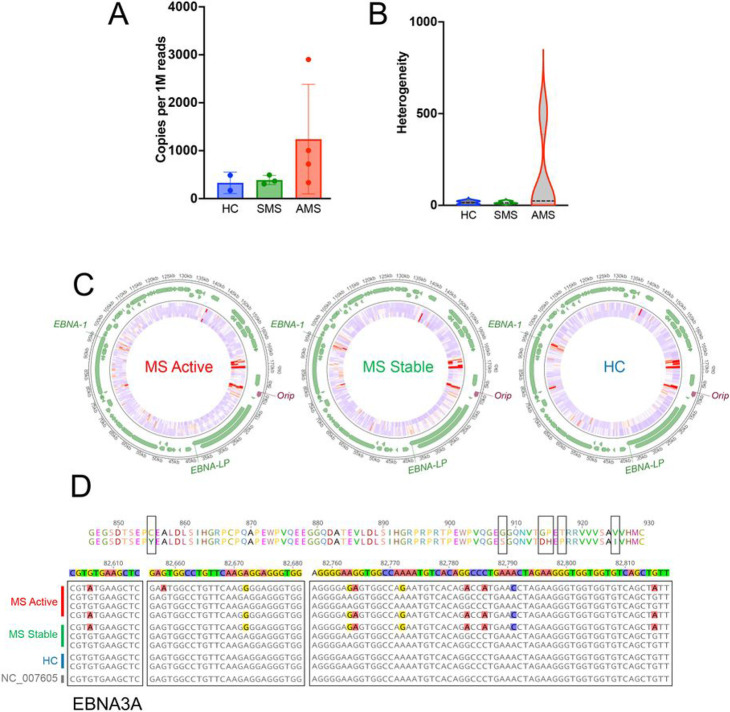
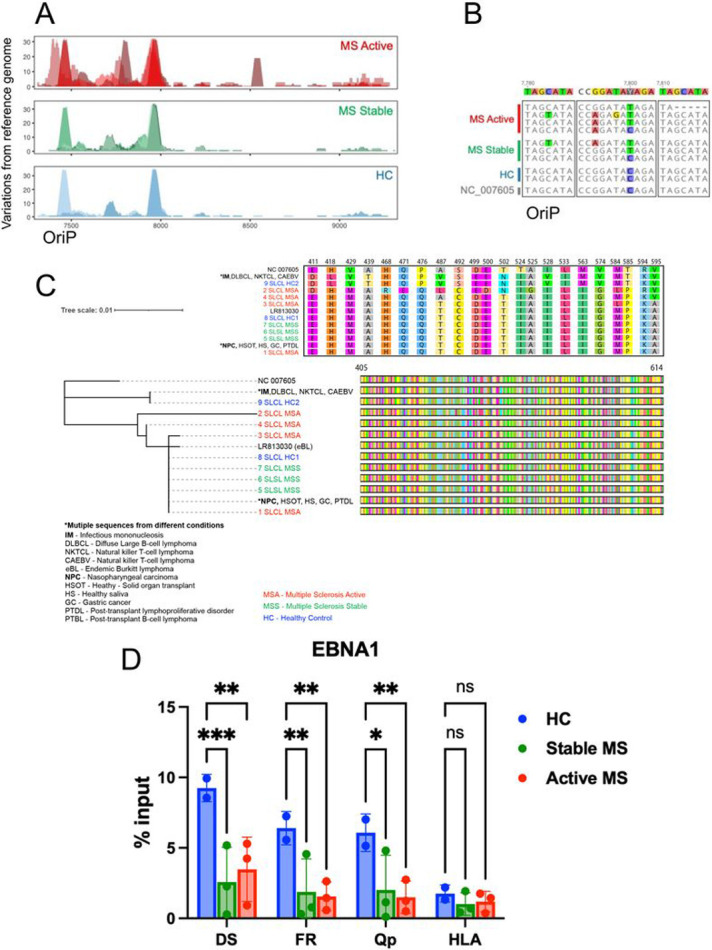
Epidemiological studies have demonstrated that Epstein-Barr virus (EBV) is a known etiologic risk factor, and perhaps prerequisite, for the development of MS. EBV establishes life-long latent infection in a subpopulation of memory B cells. Although the role of memory B cells in the pathobiology of MS is well established, studies characterizing EBV-associated mechanisms of B cell inflammation and disease pathogenesis in EBV (+) B cells from MS patients are limited. Accordingly, we analyzed spontaneous lymphoblastoid cell lines (SLCLs) from multiple sclerosis patients and healthy controls to study host-virus interactions in B cells, in the context of an individual's endogenous EBV. We identify differences in EBV gene expression and regulation of both viral and cellular genes in SLCLs. Our data suggest that EBV latency is dysregulated in MS SLCLs with increased lytic gene expression observed in MS patient B cells, especially those generated from samples obtained during "active" disease. Moreover, we show increased inflammatory gene expression and cytokine production in MS patient SLCLs and demonstrate that tenofovir alafenamide, an antiviral that targets EBV replication, decreases EBV viral loads, EBV lytic gene expression, and EBV-mediated inflammation in both SLCLs and in a mixed lymphocyte assay. Collectively, these data suggest that dysregulation of EBV latency in MS drives a pro-inflammatory, pathogenic phenotype in memory B cells and that this response can be attenuated by suppressing EBV lytic activation. This study provides further support for the development of antiviral agents that target EBV-infection for use in MS.
Keywords: Epstein-Barr Virus; Multiple Sclerosis; latency; lymphoblastoid cell lines (LCLs); lytic activation.
Conflict of interest statement
Conflict of Interest Statement PML is an advisor and has ownership interest in Vironika, LLC.
Figures


References
-
- Young L. S., Arrand J. R. & Murray P. G. in Human Herpesviruses: Biology, Therapy, and Immunoprophylaxis (eds Arvin A. et al. ) (2007). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
