

This is a preprint.
Missense mutations in CRX homeodomain cause dominant retinopathies through two distinct mechanisms
- PMID: 36778408
- PMCID: PMC9915647
- DOI: 10.1101/2023.02.01.526652
Missense mutations in CRX homeodomain cause dominant retinopathies through two distinct mechanisms
Update in
-
Missense mutations in CRX homeodomain cause dominant retinopathies through two distinct mechanisms.Elife. 2023 Nov 14;12:RP87147. doi: 10.7554/eLife.87147. Elife. 2023. PMID: 37963072 Free PMC article.
Abstract
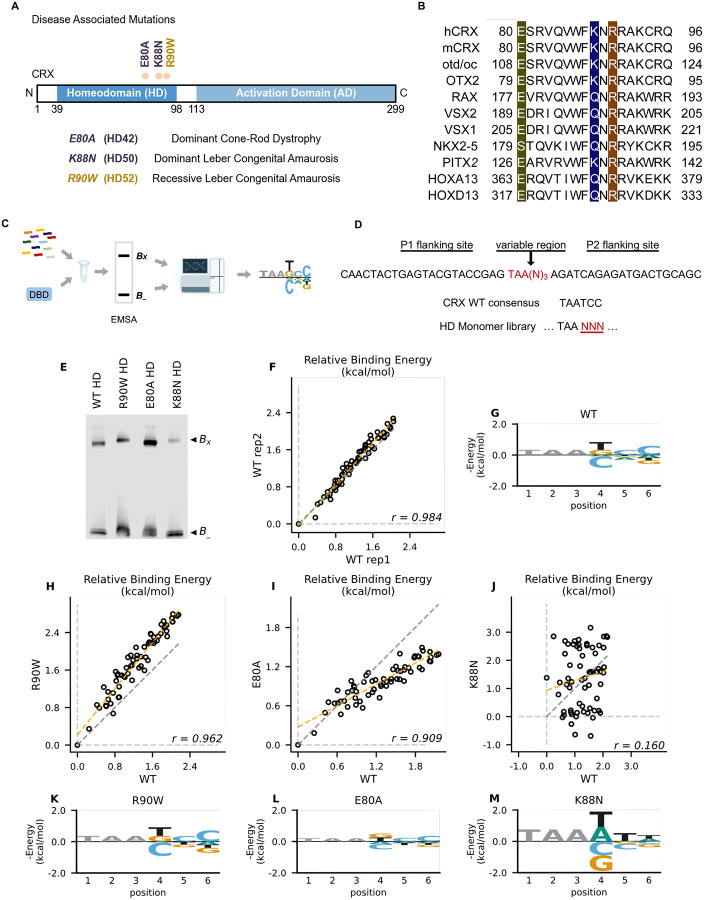
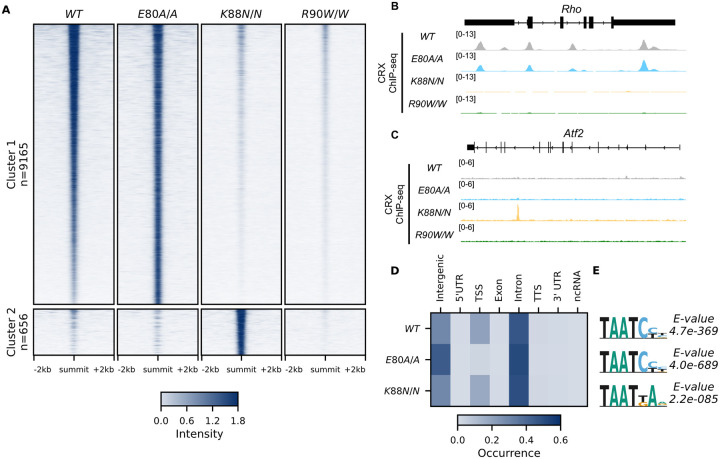
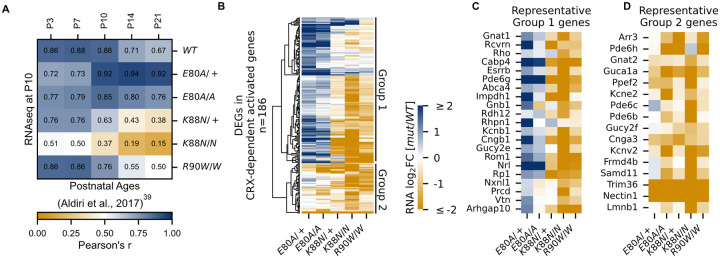
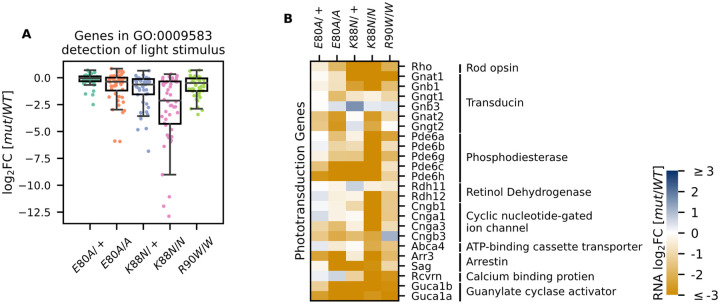
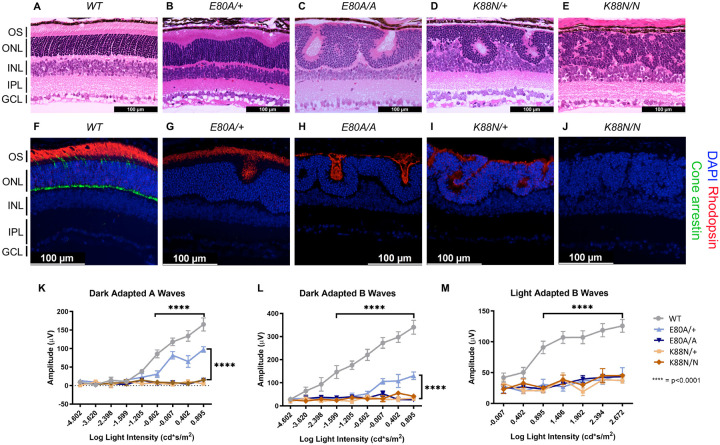
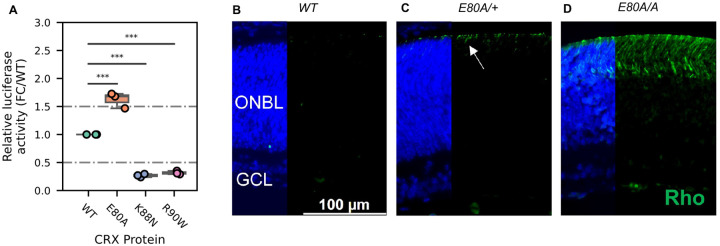
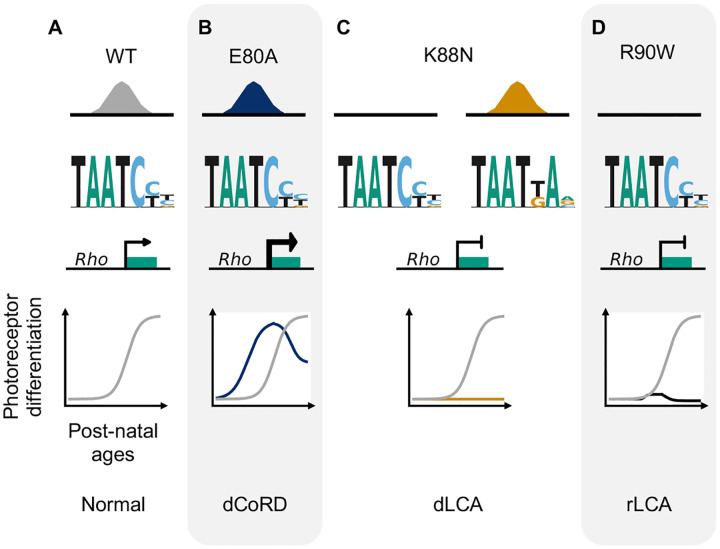
Homeodomain transcription factors (HD TFs) are instrumental to vertebrate development. Mutations in HD TFs have been linked to human diseases, but their pathogenic mechanisms remain elusive. Here we use Cone-Rod Homeobox (CRX) as a model to decipher the disease-causing mechanisms of two HD mutations, p.E80A and p.K88N, that produce severe dominant retinopathies. Through integrated analysis of molecular and functional evidence in vitro and in knock-in mouse models, we uncover two novel gain-of-function mechanisms: p.E80A increases CRX-mediated transactivation of canonical CRX target genes in developing photoreceptors; p.K88N alters CRX DNA-binding specificity resulting in binding at ectopic sites and severe perturbation of CRX target gene expression. Both mechanisms produce novel retinal morphological defects and hinder photoreceptor maturation distinct from loss-of-function models. This study reveals the distinct roles of E80 and K88 residues in CRX HD regulatory functions and emphasizes the importance of transcriptional precision in normal development.
Keywords: CRX mutations; DNA binding; Homeodomain; gene expression; inherited retinal disease; knock-in mouse models; photoreceptor development; transcription factor.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Mark M., Rijli F.M. & Chambon P. Homeobox Genes in Embryogenesis and Pathogenesis. Pediatric Research 42, 421–429 (1997). - PubMed
-
- Lewis E.B. A gene complex controlling segmentation in Drosophila. Nature 276, 565–570 (1978). - PubMed
-
- Zagozewski J.L., Zhang Q., Pinto V.I., Wigle J.T. & Eisenstat D.D. The role of homeobox genes in retinal development and disease. Developmental Biology 393, 195–208 (2014). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials