

This is a preprint.
Eukaryotic-like gephyrin and cognate membrane receptor coordinate corynebacterial cell division and polar elongation
- PMID: 36778425
- PMCID: PMC9915583
- DOI: 10.1101/2023.02.01.526586
Eukaryotic-like gephyrin and cognate membrane receptor coordinate corynebacterial cell division and polar elongation
Update in
-
Eukaryotic-like gephyrin and cognate membrane receptor coordinate corynebacterial cell division and polar elongation.Nat Microbiol. 2023 Oct;8(10):1896-1910. doi: 10.1038/s41564-023-01473-0. Epub 2023 Sep 7. Nat Microbiol. 2023. PMID: 37679597 Free PMC article.
Abstract
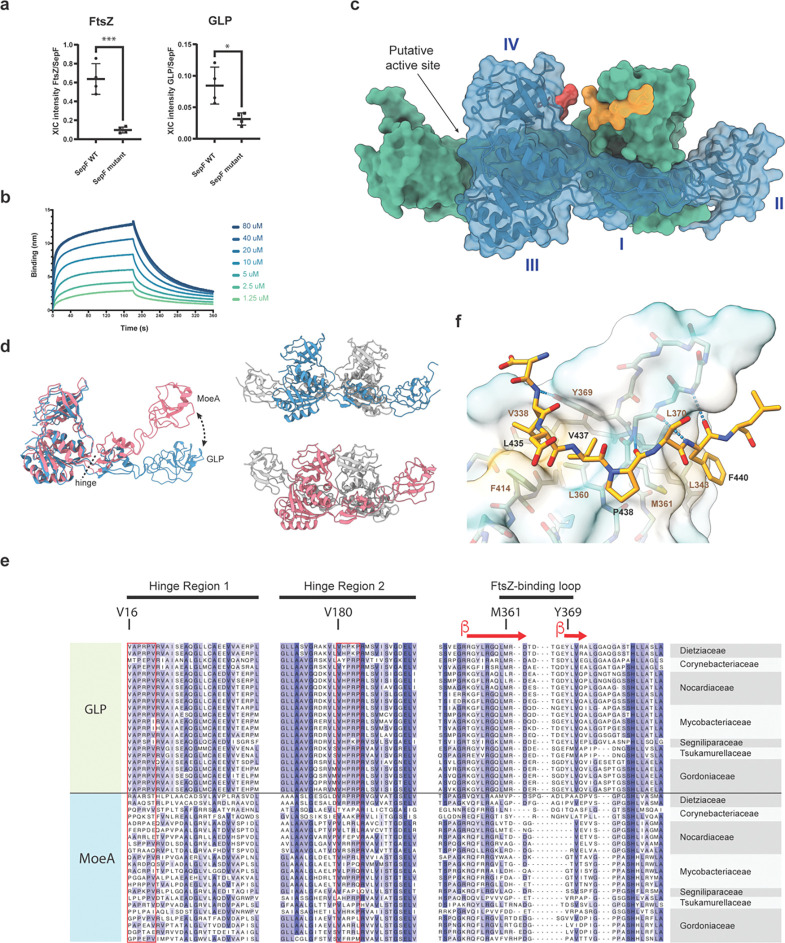
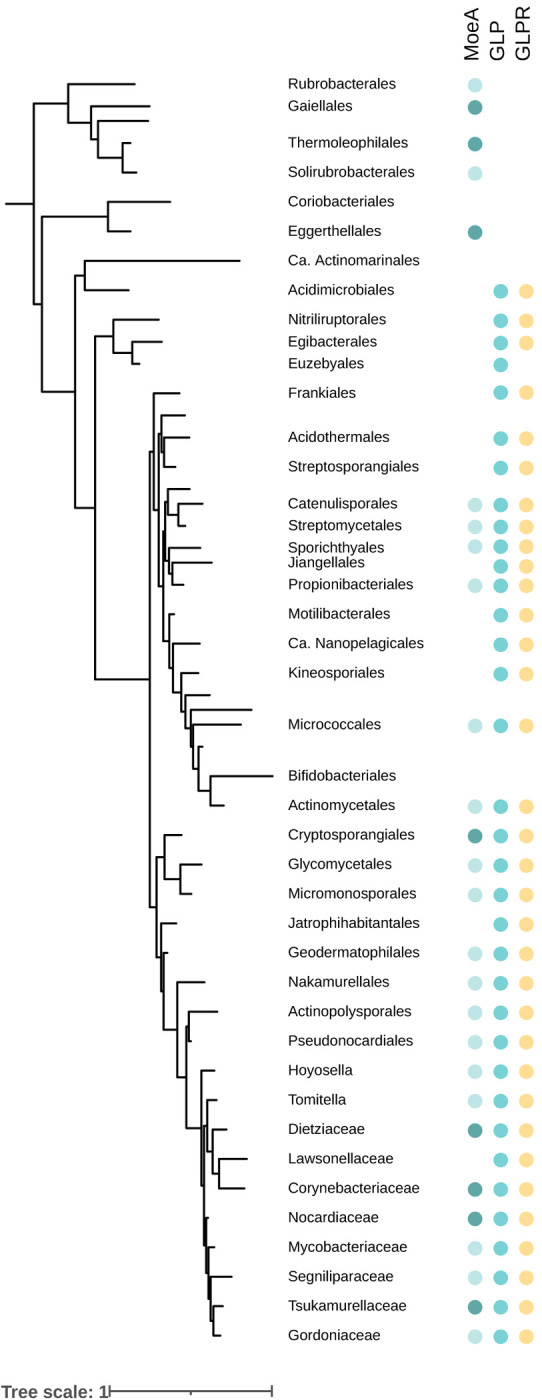
The order Corynebacteriales includes major industrial and pathogenic actinobacteria such as Corynebacterium glutamicum or Mycobacterium tuberculosis . Their elaborate multi-layered cell wall, composed primarily of the mycolyl-arabinogalactan-peptidoglycan complex, and their polar growth mode impose a stringent coordination between the septal divisome, organized around the tubulin-like protein FtsZ, and the polar elongasome, assembled around the tropomyosin-like protein Wag31. Here, we report the identification of two new divisome members, a gephyrin-like repurposed molybdotransferase (GLP) and its membrane receptor (GLPR). We show that the interplay between the GLPR/GLP module, FtsZ and Wag31 is crucial for orchestrating cell cycle progression. Our results provide a detailed molecular understanding of the crosstalk between two essential machineries, the divisome and elongasome, and reveal that Corynebacteriales have evolved a protein scaffold to control cell division and morphogenesis similar to the gephyrin/GlyR system that in higher eukaryotes mediates synaptic signaling through network organization of membrane receptors and the microtubule cytoskeleton.
Conflict of interest statement
Competing interests
The authors declare no competing financial interests.
Figures




References
-
- Hirota H., Ryter A., and Jacob F. (1968). Thermosensitive Mutants of E. coli Affected in the Processes of DNA Synthesis and Cellular Division. Cold Spring Harb Symp Quant Biol 33, 677–693. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources