

Single Particle Analysis of H3N2 Influenza Entry Differentiates the Impact of the Sialic Acids (Neu5Ac and Neu5Gc) on Virus Binding and Membrane Fusion
- PMID: 36779754
- PMCID: PMC10062150
- DOI: 10.1128/jvi.01463-22
Single Particle Analysis of H3N2 Influenza Entry Differentiates the Impact of the Sialic Acids (Neu5Ac and Neu5Gc) on Virus Binding and Membrane Fusion
Abstract
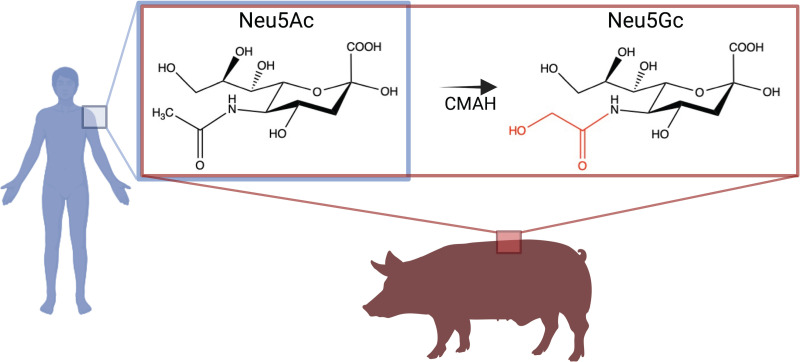
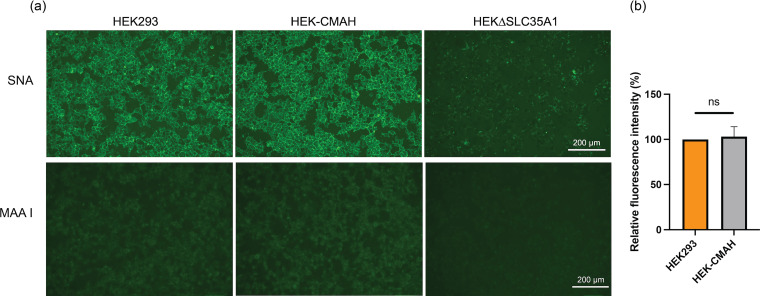
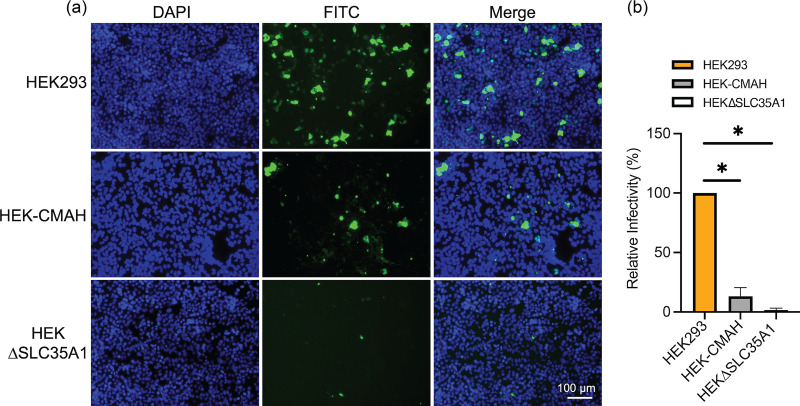
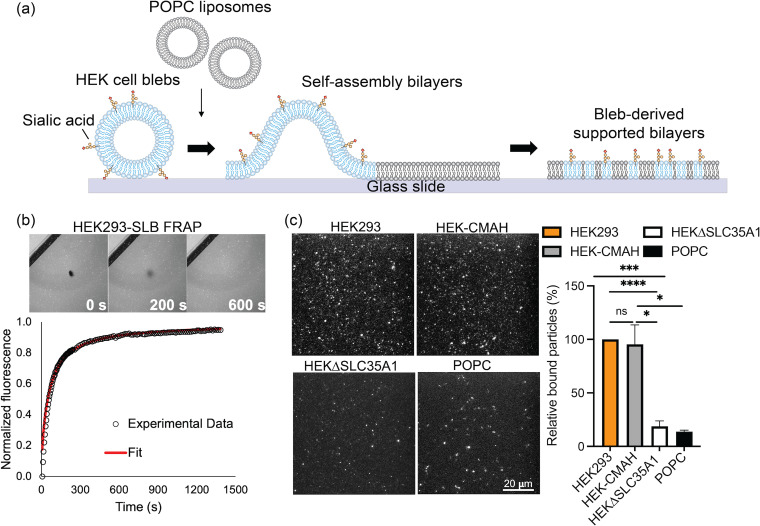
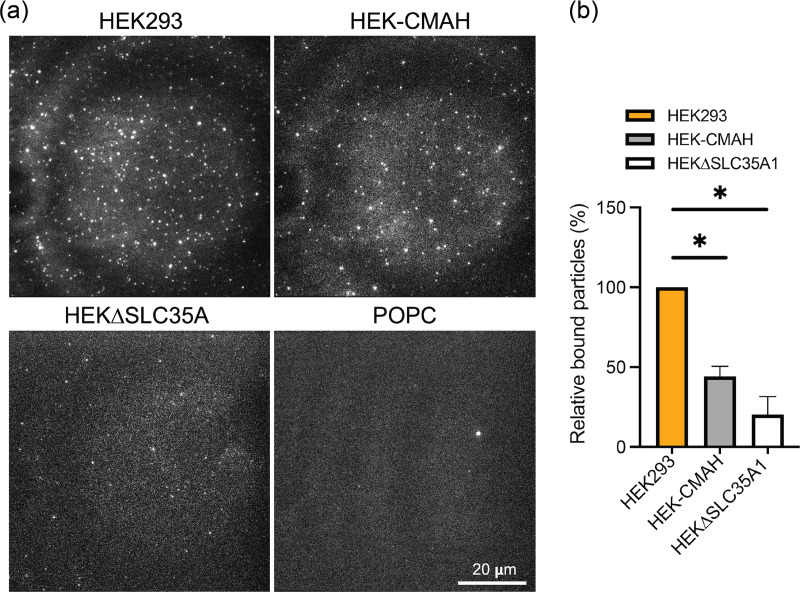
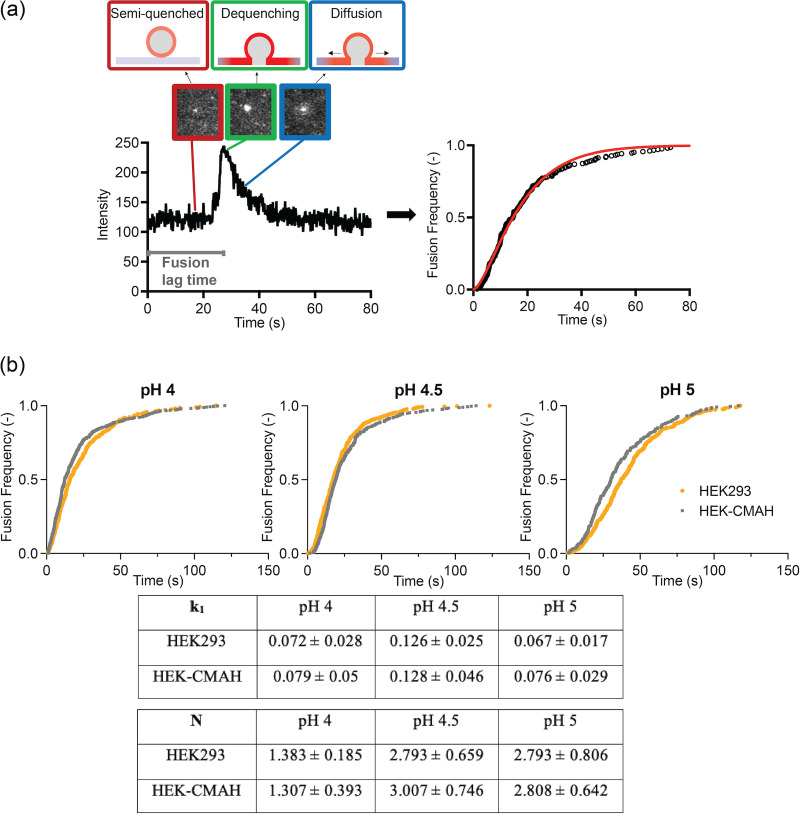
Entry of influenza A viruses (IAVs) into host cells is initiated by binding to sialic acids (Sias), their primary host cell receptor, followed by endocytosis and membrane fusion to release the viral genome into the cytoplasm of the host cell. Host tropism is affected by these entry processes, with a primary factor being receptor specificity. Sias exist in several different chemical forms, including the hydroxylated N-glycolylneuraminic acid (Neu5Gc), which is found in many hosts; however, it has not been clear how modified Sias affect viral binding and entry. Neu5Gc is commonly found in many natural influenza hosts, including pigs and horses, but not in humans or ferrets. Here, we engineered HEK293 cells to express the hydoxylase gene (CMAH) that converts Neu5Ac to Neu5Gc, or knocked out the Sia-CMP transport gene (SLC35A1), resulting in cells that express 95% Neu5Gc or minimal level of Sias, respectively. H3N2 (X-31) showed significantly reduced infectivity in Neu5Gc-rich cells compared to wild-type HEK293 (>95% Neu5Ac). To determine the effects on binding and fusion, we generated supported lipid bilayers (SLBs) derived from the plasma membranes of these cells and carried out single particle microscopy. H3N2 (X-31) exhibited decreased binding to Neu5Gc-containing SLBs, but no significant difference in H3N2 (X-31)'s fusion kinetics to either SLB type, suggesting that reduced receptor binding does not affect subsequent membrane fusion. This finding suggests that for this virus to adapt to host cells rich in Neu5Gc, only receptor affinity changes are required without further adaptation of virus fusion machinery. IMPORTANCE Influenza A virus (IAV) infections continue to threaten human health, causing over 300,000 deaths yearly. IAV infection is initiated by the binding of influenza glycoprotein hemagglutinin (HA) to host cell sialic acids (Sias) and the subsequent viral-host membrane fusion. Generally, human IAVs preferentially bind to the Sia N-acetylneuraminic acid (Neu5Ac). Yet, other mammalian hosts, including pigs, express diverse nonhuman Sias, including N-glycolylneuraminic acid (Neu5Gc). The role of Neu5Gc in human IAV infections in those hosts is not well-understood, and the variant form may play a role in incidents of cross-species transmission and emergence of new epidemic variants. Therefore, it is important to investigate how human IAVs interact with Neu5Ac and Neu5Gc. Here, we use membrane platforms that mimic the host cell surface to examine receptor binding and membrane fusion events of human IAV H3N2. Our findings improve the understanding of viral entry mechanisms that can affect host tropism and virus evolution.
Keywords: Neu5Ac; Neu5Gc; fusion kinetics; influenza; membrane fusion; membranes; sialic acid; single particle tracking; supported lipid bilayer; tropism; virus entry.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Lewis AL, Chen X, Schnaar RL, et al. 2022. Sialic acids and other nonulosonic acids. In Varki A, Cummings RD, Esko JD, et al. Essentials of glycobiology [Internet]. 4th ed. Cold spring harbor (NY): Cold spring harbor laboratory press. Chapter 15. https://www.ncbi.nlm.nih.gov/books/NBK579976/.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
