

Interleukin-23 Regulates Inflammatory Osteoclastogenesis via Activation of CLEC5A(+) Osteoclast Precursors
- PMID: 36787107
- PMCID: PMC10423744
- DOI: 10.1002/art.42478
Interleukin-23 Regulates Inflammatory Osteoclastogenesis via Activation of CLEC5A(+) Osteoclast Precursors
Abstract
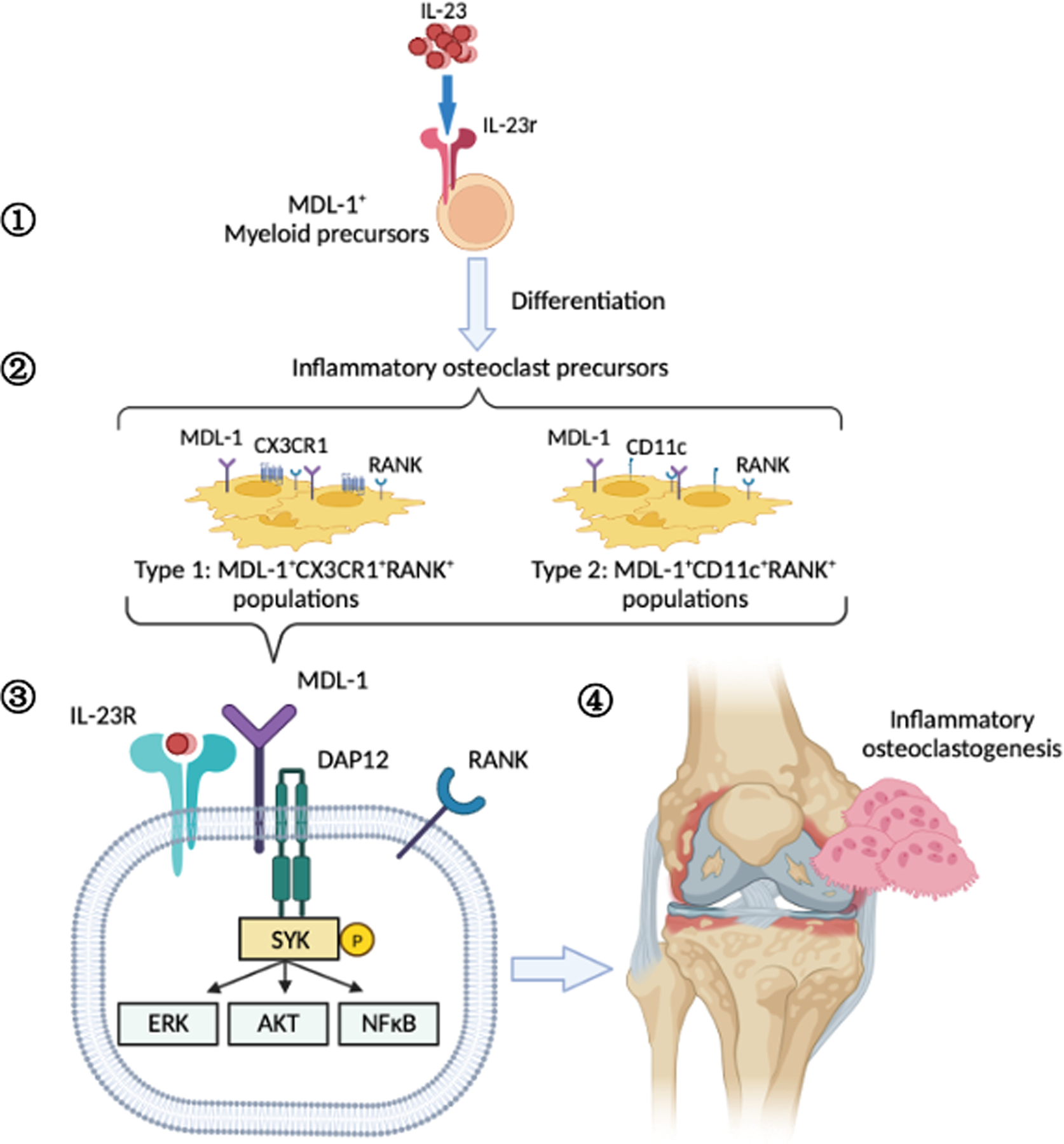
Objective: To investigate the role of interleukin-23 (IL-23) in pathologic bone remodeling in inflammatory arthritis.
Methods: In this study we investigated the role of IL-23 in osteoclast differentiation and activation using in vivo gene transfer techniques in wild-type and myeloid DNAX-activation protein 12-associating lectin-1 (MDL-1)-deficient mice, and by performing in vitro and in vivo osteoclastogenesis assays using spectral flow cytometry, micro-computed tomography analysis, Western blotting, and immunoprecipitation.
Results: Herein, we show that IL-23 induces the expansion of a myeloid osteoclast precursor population and supports osteoclastogenesis and bone resorption in inflammatory arthritis. Genetic ablation of C-type lectin domain family member 5A, also known as MDL-1, prevents the induction of osteoclast precursors by IL-23 that is associated with bone destruction, as commonly observed in inflammatory arthritis. Moreover, osteoclasts derived from the bone marrow of MDL-1-deficient mice showed impaired osteoclastogenesis, and MDL-1-/- mice had increased bone mineral density.
Conclusion: Our data show that IL-23 signaling regulates the availability of osteoclast precursors in inflammatory arthritis that could be effectively targeted for the treatment of inflammatory bone loss in inflammatory arthritis.
© 2023 American College of Rheumatology.
Conflict of interest statement
Disclosures
The authors have no financial conflicts of interest.
Figures
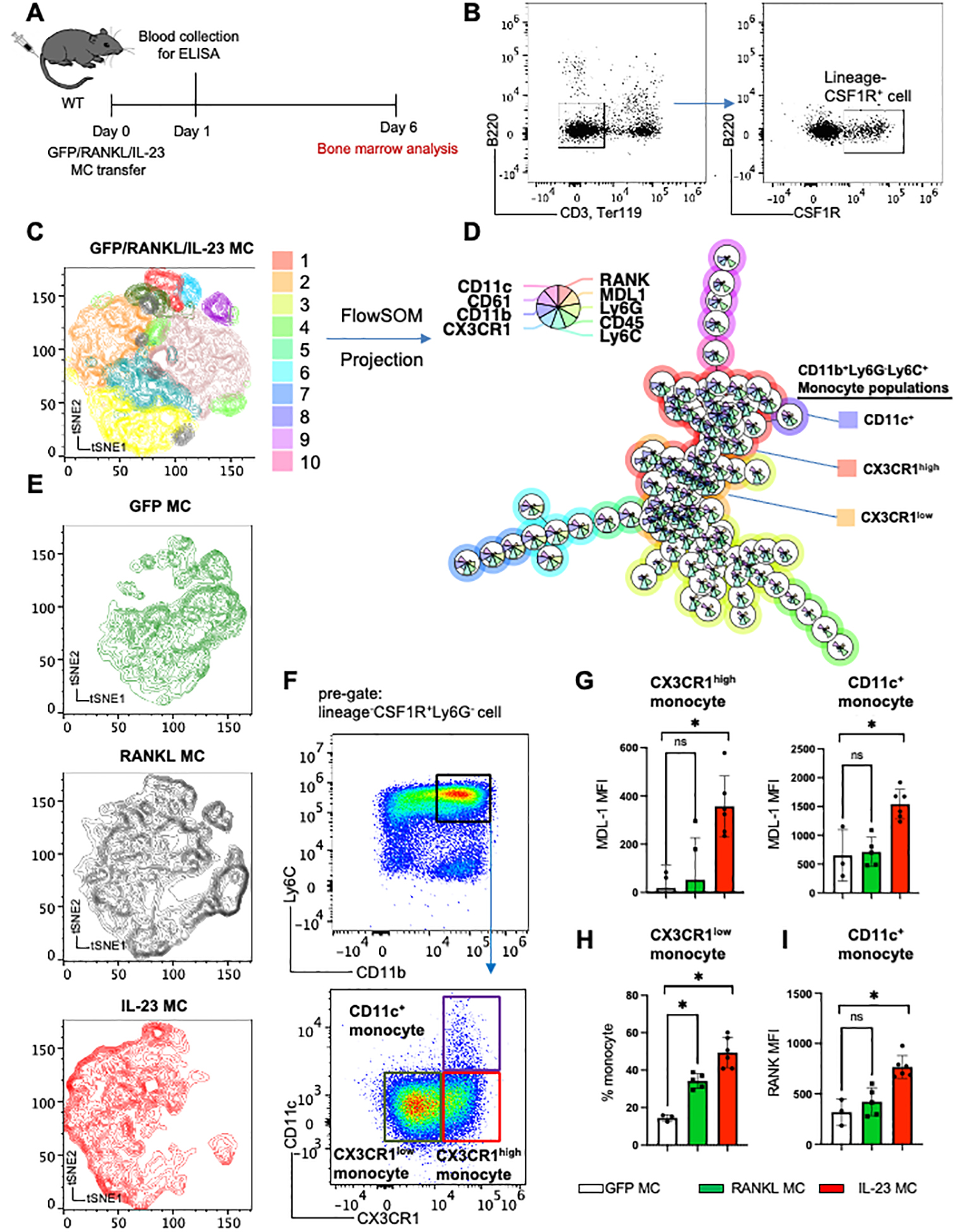
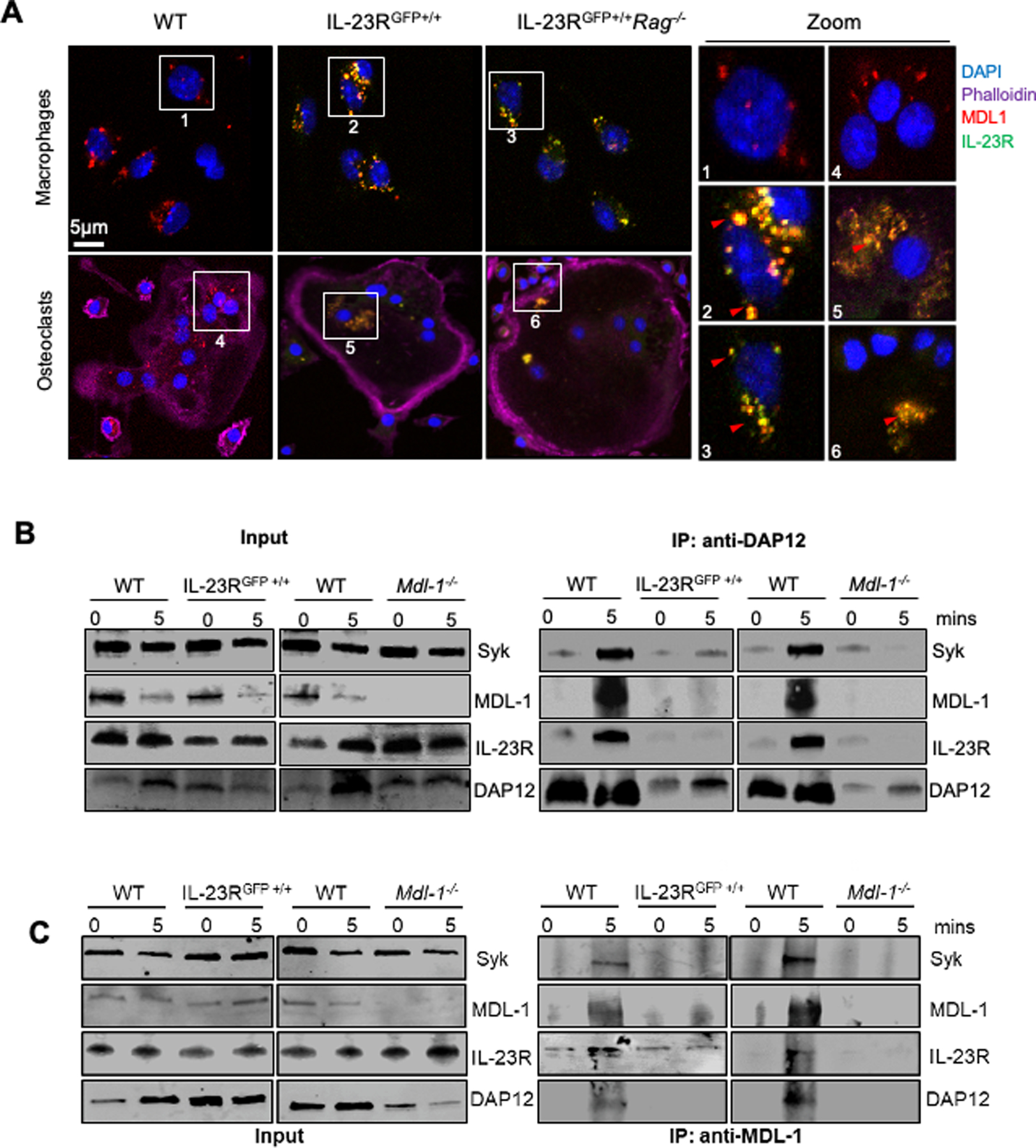

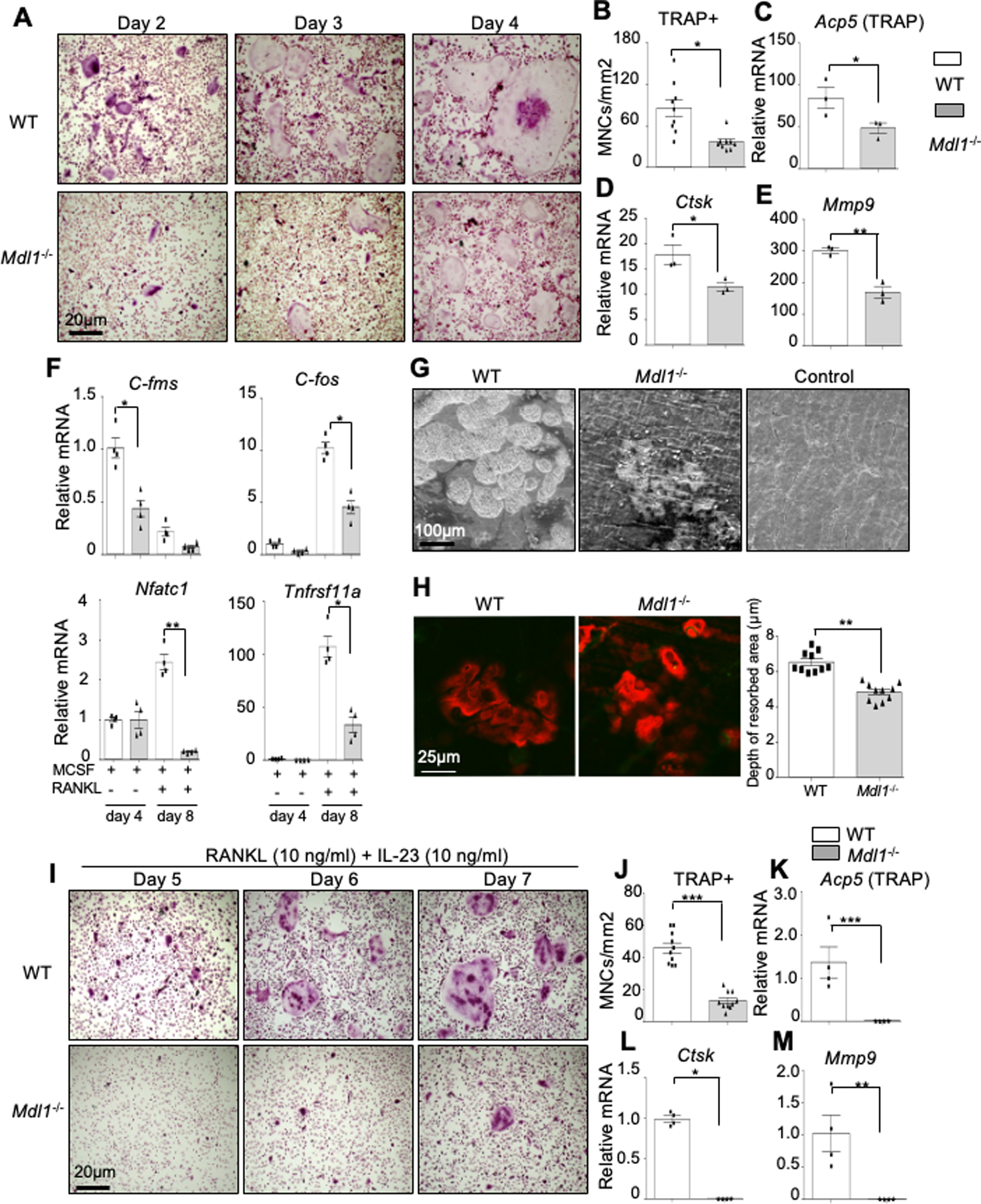
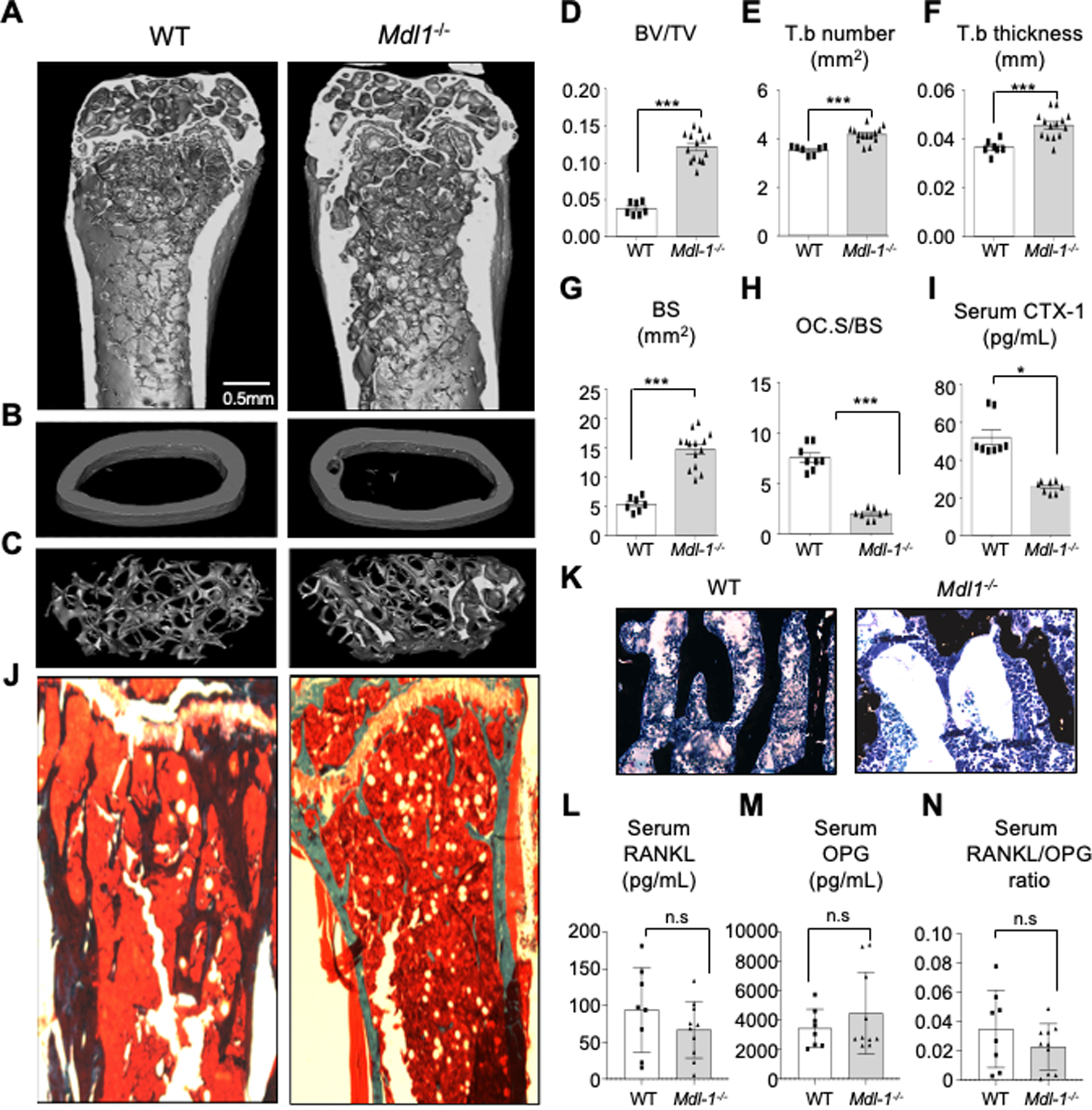
References
-
- Boyle WJ, Simonet WS, Lacey DL. Osteoclast differentiation and activation. Nature.2003;423(6937):337–42. - PubMed
-
- Koga T, Inui M, Inoue K, Kim S, Suematsu A, Kobayashi E, et al. Costimulatory signals mediated by the ITAM motif cooperate with RANKL for bone homeostasis. Nature. 2004;428(6984):758–63. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
