

Brief synaptic inhibition persistently interrupts firing of fast-spiking interneurons
- PMID: 36787751
- PMCID: PMC10121938
- DOI: 10.1016/j.neuron.2023.01.017
Brief synaptic inhibition persistently interrupts firing of fast-spiking interneurons
Abstract
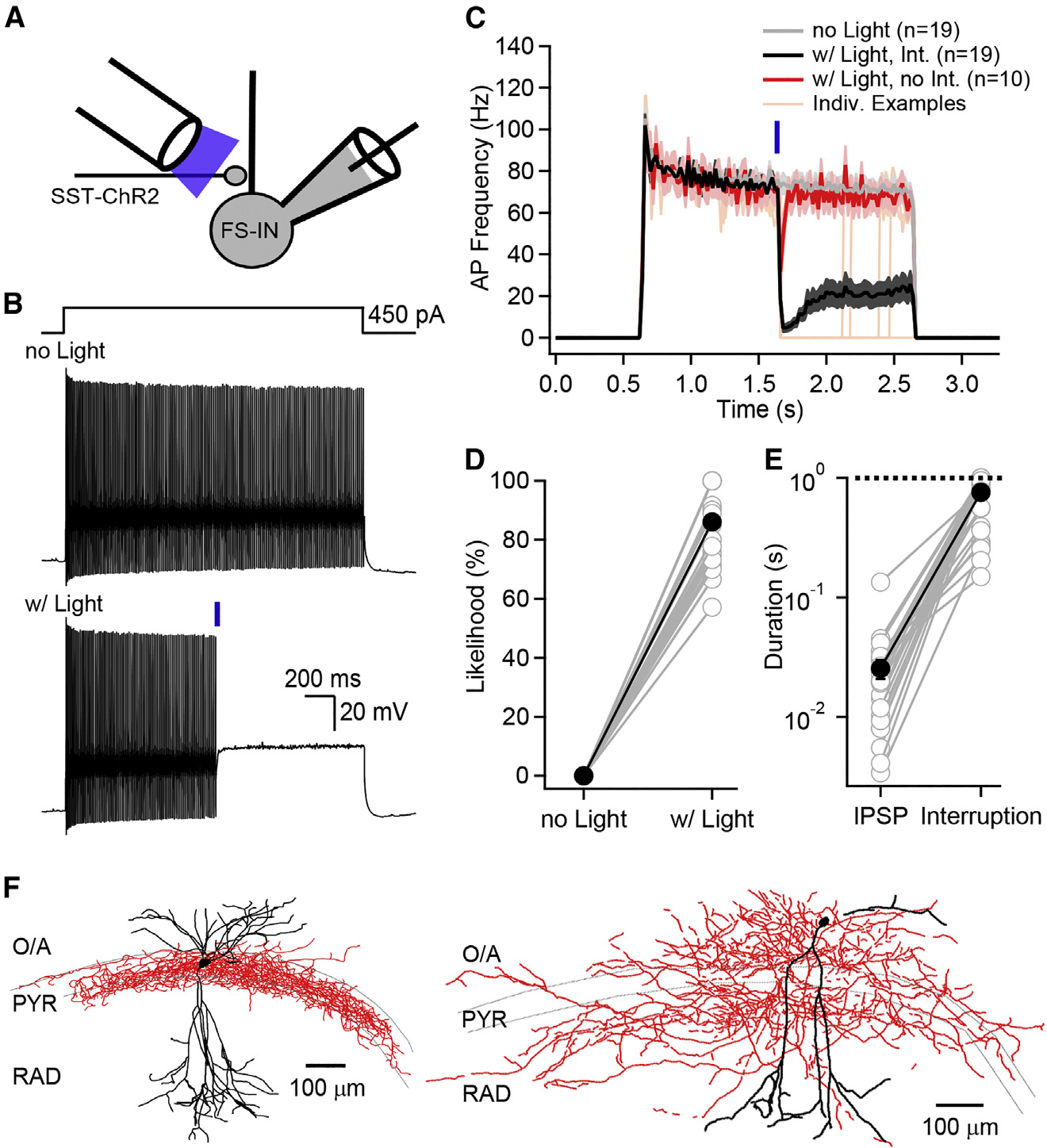
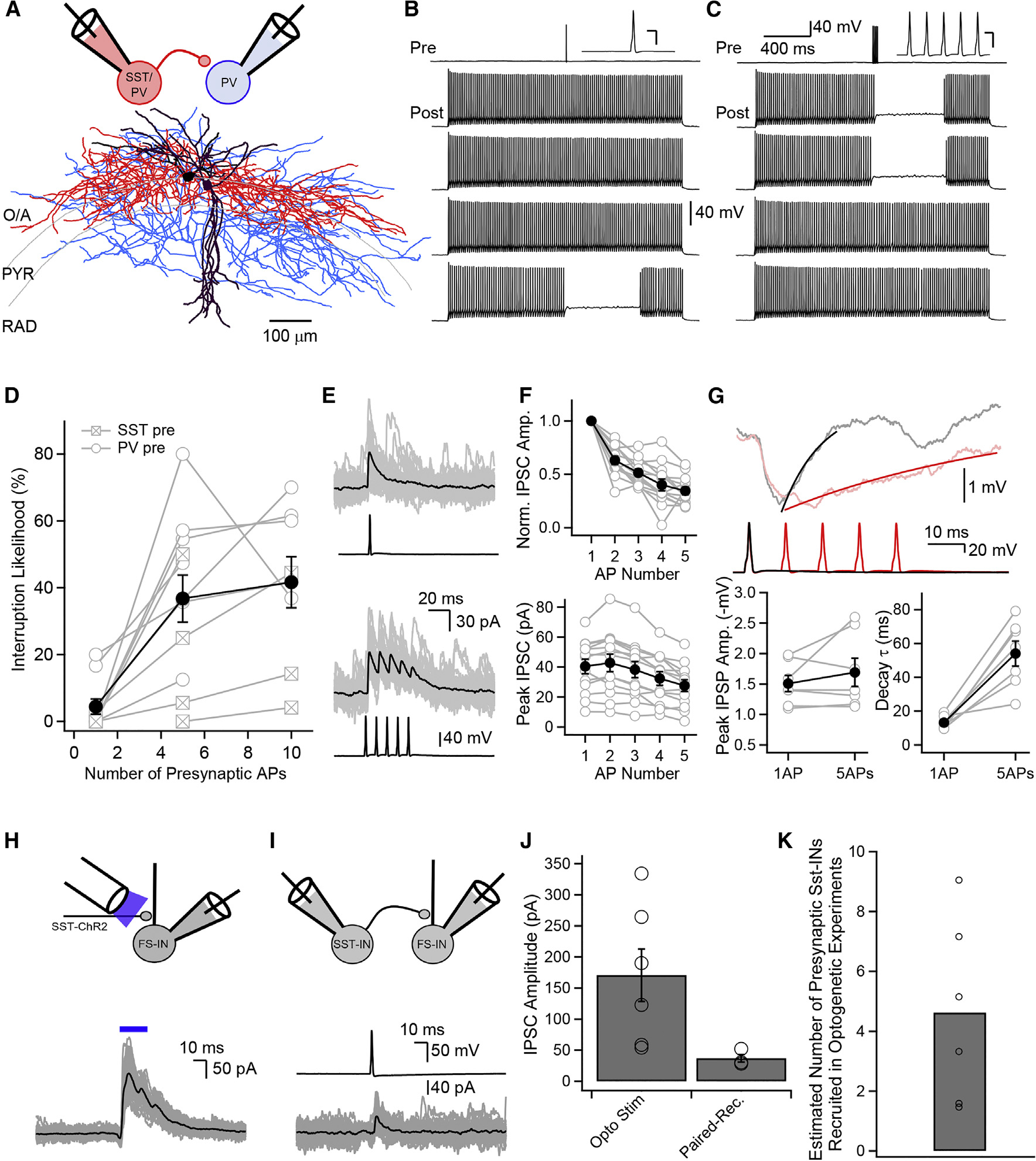
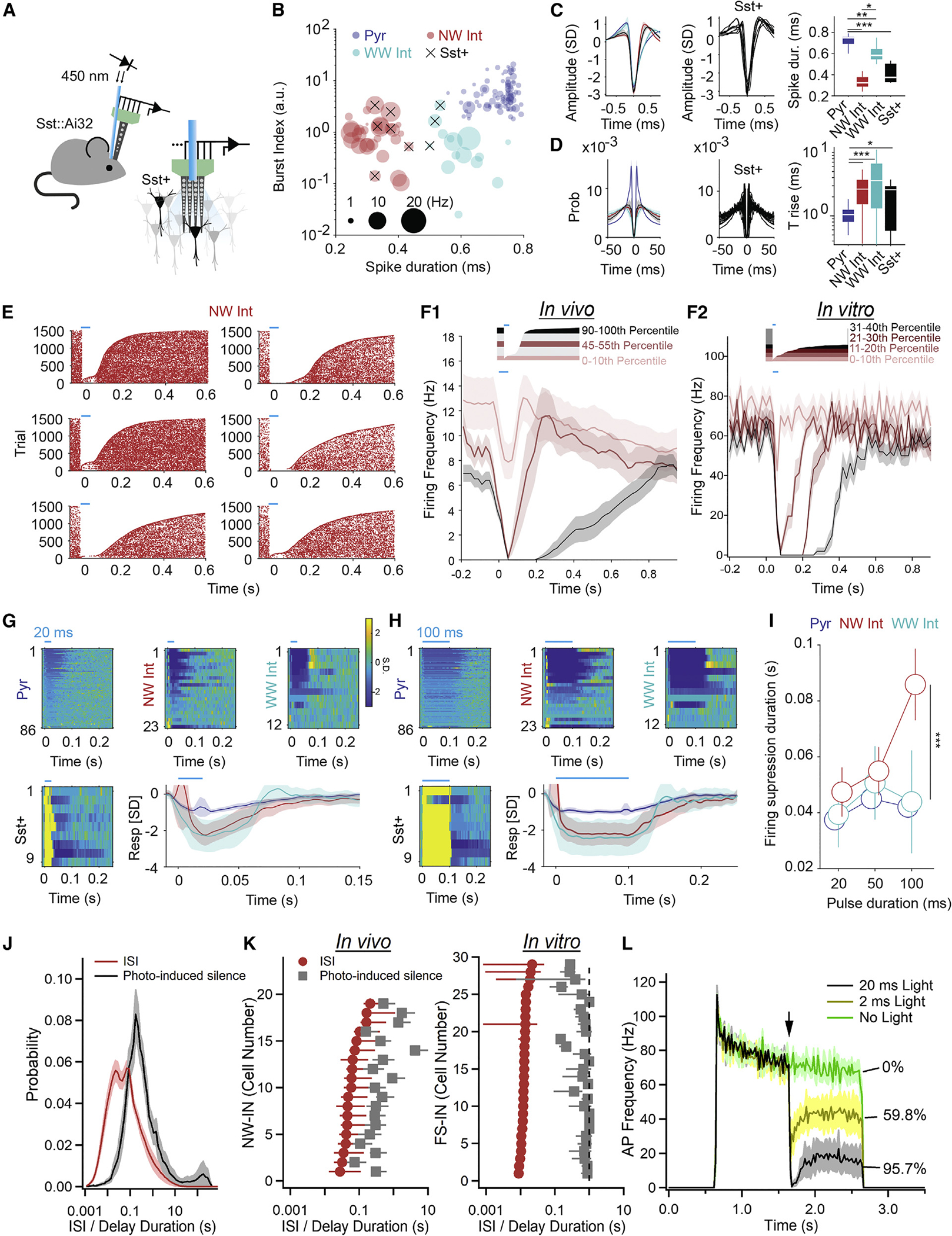
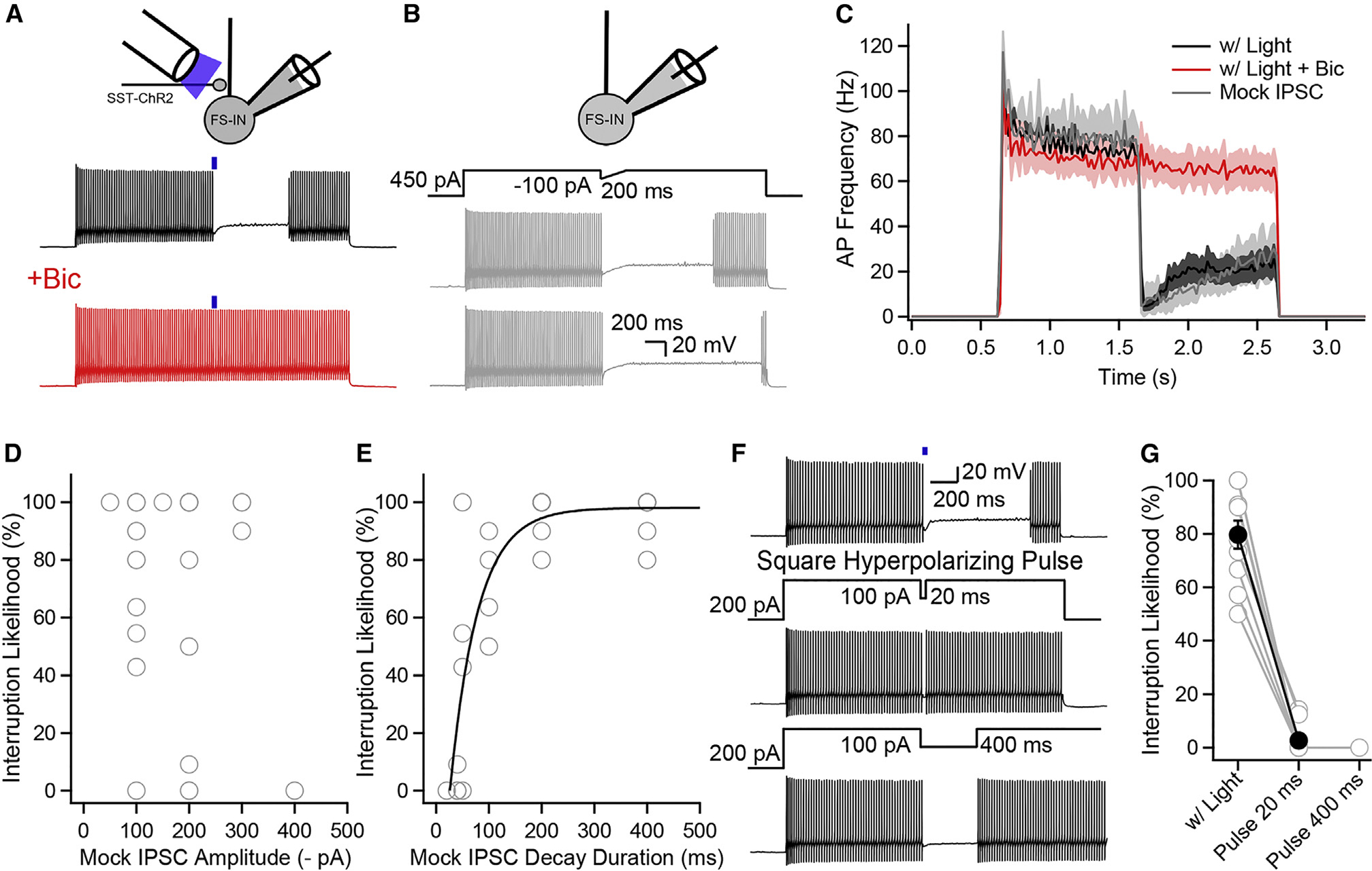
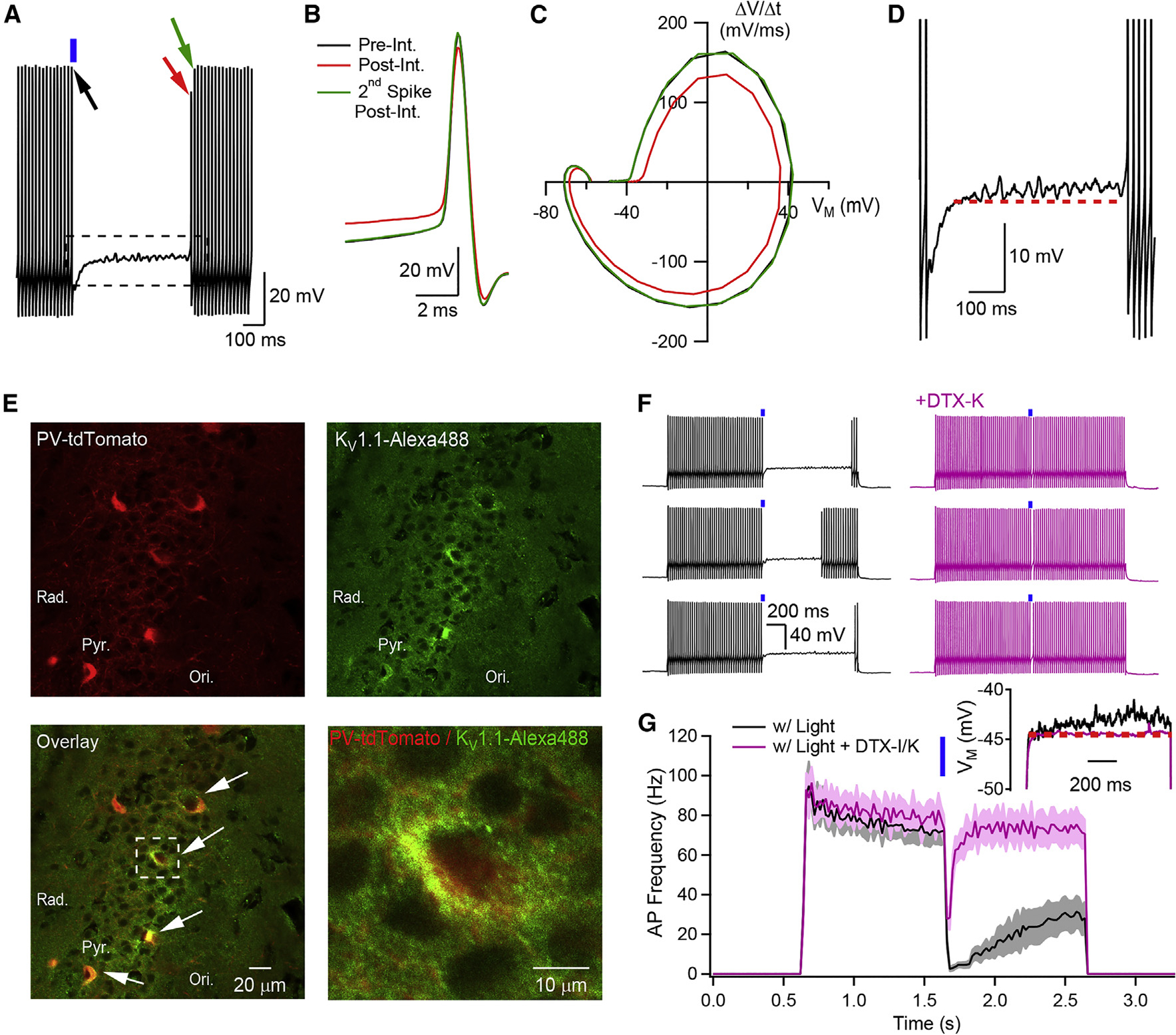
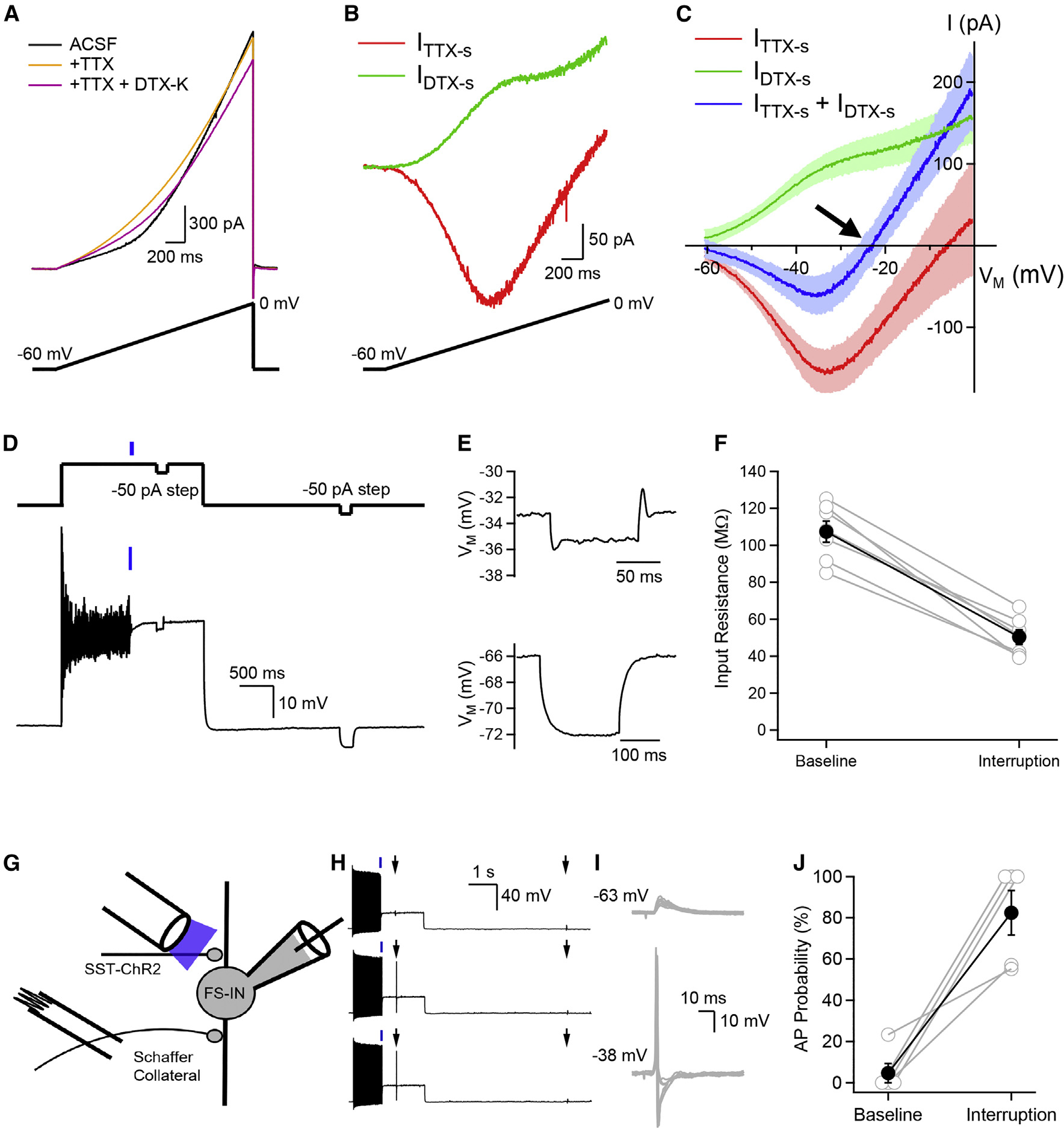
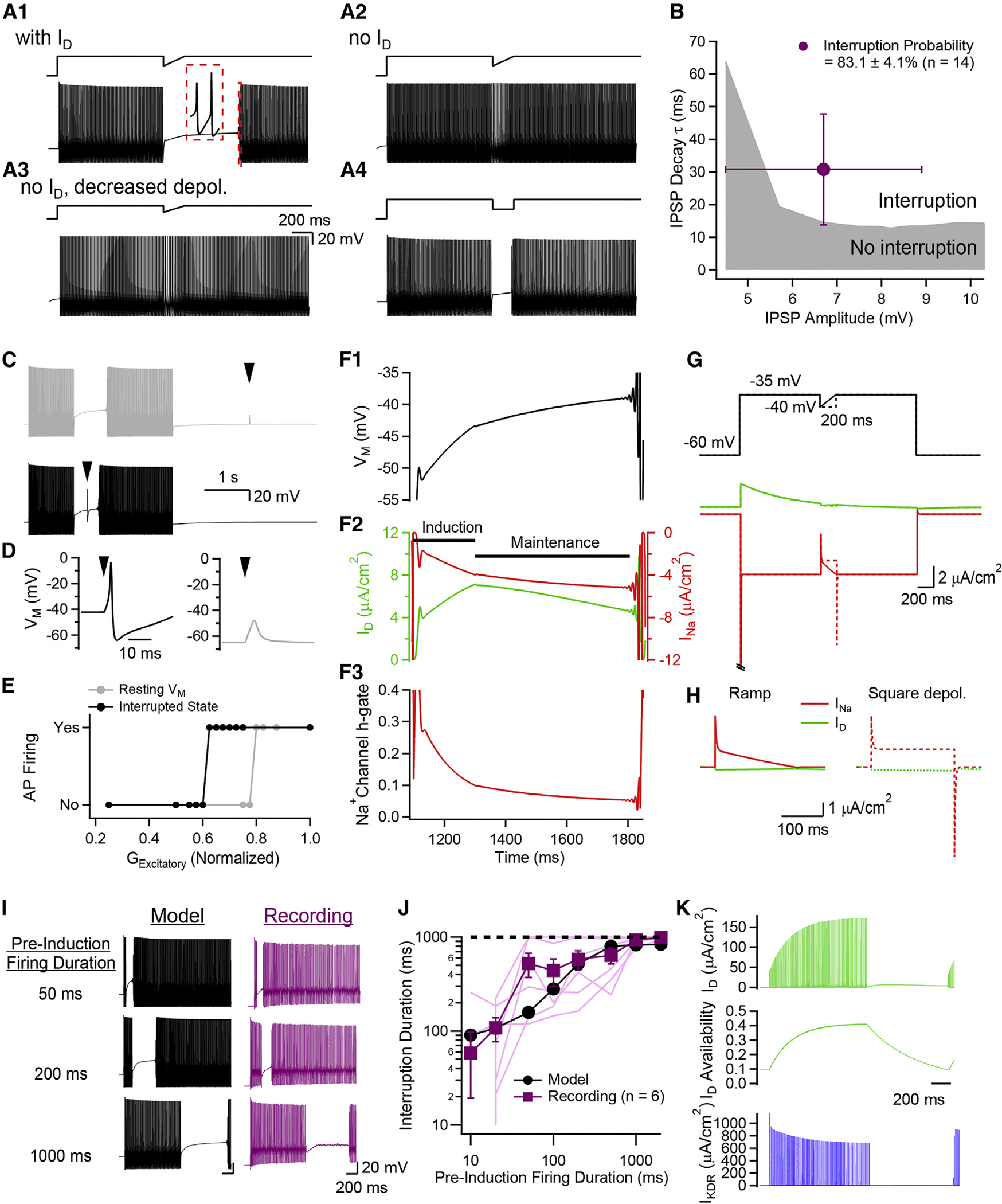
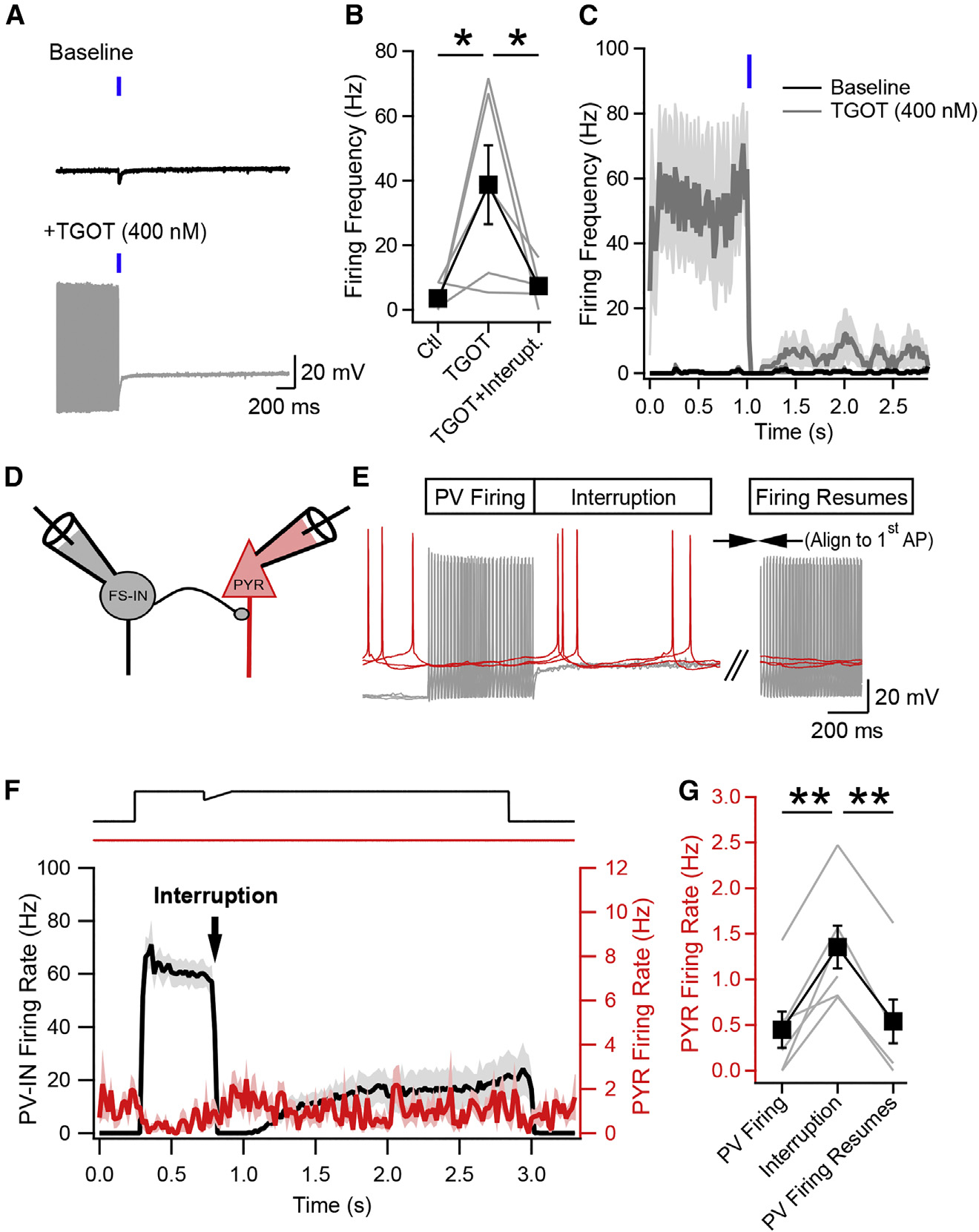
Neurons perform input-output operations that integrate synaptic inputs with intrinsic electrical properties; these operations are generally constrained by the brevity of synaptic events. Here, we report that sustained firing of CA1 hippocampal fast-spiking parvalbumin-expressing interneurons (PV-INs) can be persistently interrupted for several hundred milliseconds following brief GABAAR-mediated inhibition in vitro and in vivo. A single presynaptic neuron could interrupt PV-IN firing, occasionally with a single action potential (AP), and reliably with AP bursts. Experiments and computational modeling reveal that the persistent interruption of firing maintains neurons in a depolarized, quiescent state through a cell-autonomous mechanism. Interrupted PV-INs are strikingly responsive to Schaffer collateral inputs. The persistent interruption of firing provides a disinhibitory circuit mechanism favoring spike generation in CA1 pyramidal cells. Overall, our results demonstrate that neuronal silencing can far outlast brief synaptic inhibition owing to the well-tuned interplay between neurotransmitter release and postsynaptic membrane dynamics, a phenomenon impacting microcircuit function.
Keywords: fast-spiking interneurons; hippocampus; inhibition; persistent activity.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
