

Cross-species activation of hydrogen cyanide production by a promiscuous quorum-sensing receptor promotes Chromobacterium subtsugae competition in a dual-species model
- PMID: 36790401
- PMCID: PMC10197875
- DOI: 10.1099/mic.0.001294
Cross-species activation of hydrogen cyanide production by a promiscuous quorum-sensing receptor promotes Chromobacterium subtsugae competition in a dual-species model
Abstract
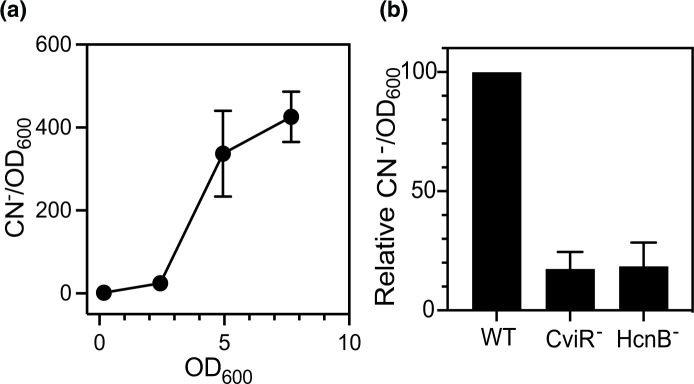
Many saprophytic bacteria have LuxR-I-type acyl-homoserine lactone (AHL) quorum-sensing systems that may be important for competing with other bacteria in complex soil communities. LuxR AHL receptors specifically interact with cognate AHLs to cause changes in expression of target genes. Some LuxR-type AHL receptors have relaxed specificity and are responsive to non-cognate AHLs. These promiscuous receptors might be used to sense and respond to AHLs produced by other bacteria by eavesdropping. We are interested in understanding the role of eavesdropping during interspecies competition. The soil saprophyte Chromobacterium subtsugae has a single AHL circuit, CviR-I, which produces and responds to N-hexanoyl-HSL (C6-HSL). The AHL receptor CviR can respond to a variety of AHLs in addition to C6-HSL. In prior studies we have utilized a coculture model with C. subtsugae and another soil saprophyte, Burkholderia thailandensis. Using this model, we previously showed that promiscuous activation of CviR by B. thailandensis AHLs provides a competitive advantage to C. subtsugae. Here, we show that B. thailandensis AHLs activate transcription of dozens of genes in C. subtsugae, including the hcnABC genes coding for production of hydrogen cyanide. We show that hydrogen cyanide production is population density-dependent and demonstrate that the cross-induction of hydrogen cyanide by B. thailandensis AHLs provides a competitive advantage to C. subtsugae. Our results provide new information on C. subtsugae quorum sensing and are the basis for future studies aimed at understanding the role of eavesdropping in interspecies competition.
Keywords: Chromobacterium; antibiotic; coculture; competition; cyanide; quorum sensing.
Conflict of interest statement
The authors declare that there are no conflicts of interest.
Figures




References
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources