

Phase separation of an actin nucleator by junctional microtubules regulates epithelial function
- PMID: 36791197
- PMCID: PMC9931218
- DOI: 10.1126/sciadv.adf6358
Phase separation of an actin nucleator by junctional microtubules regulates epithelial function
Abstract
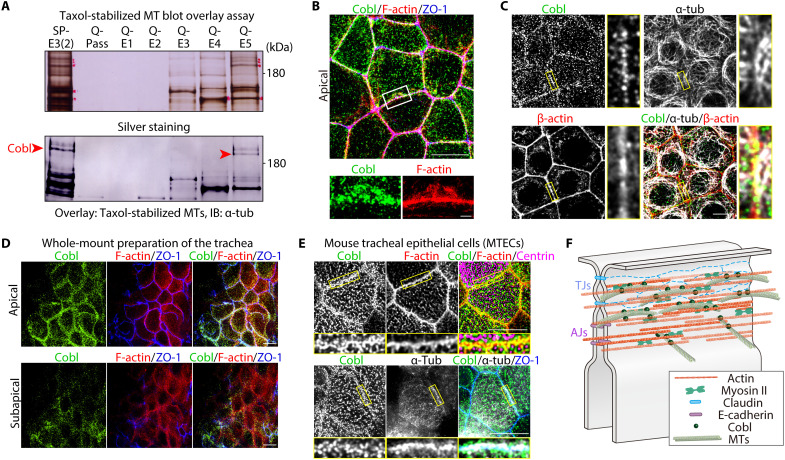
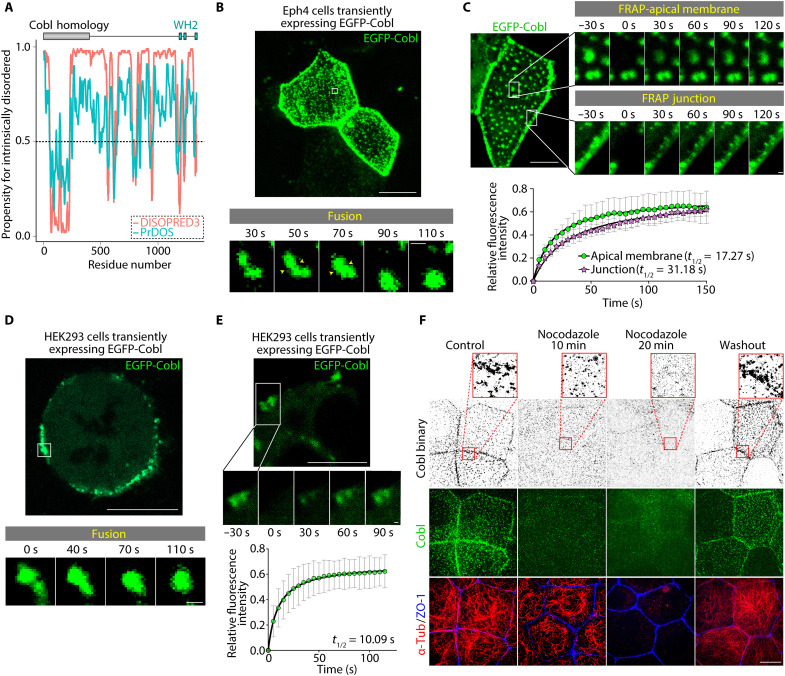
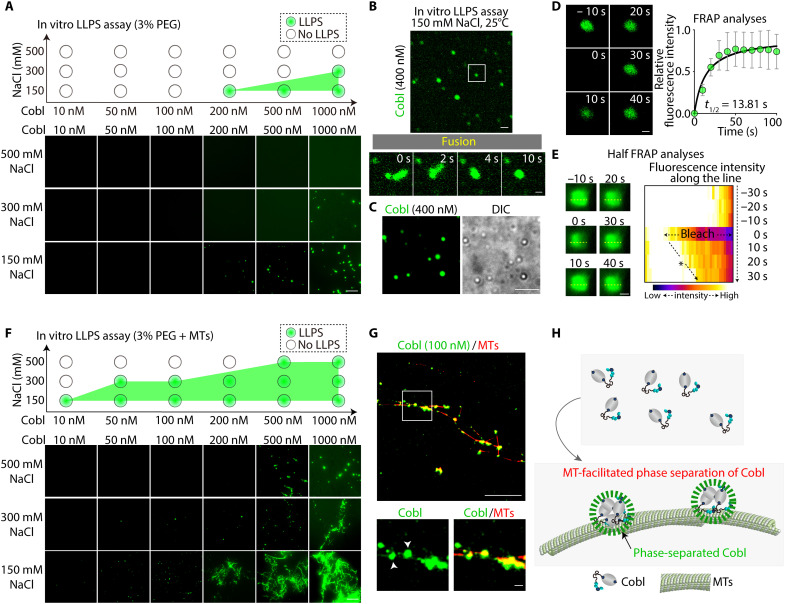
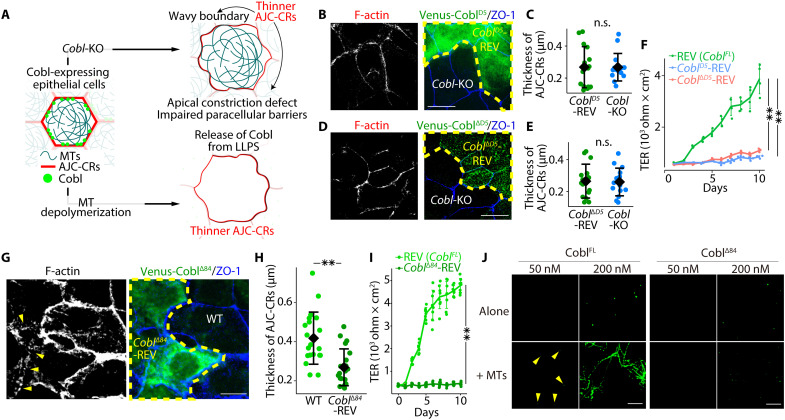
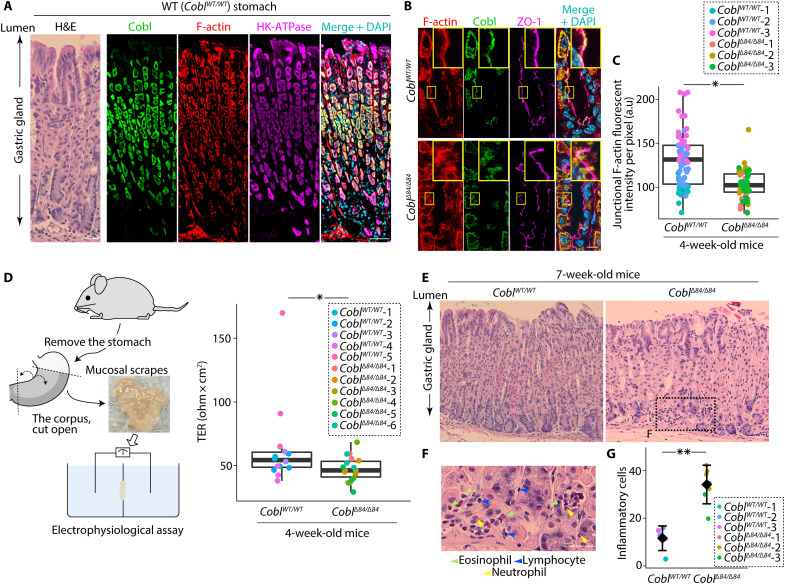
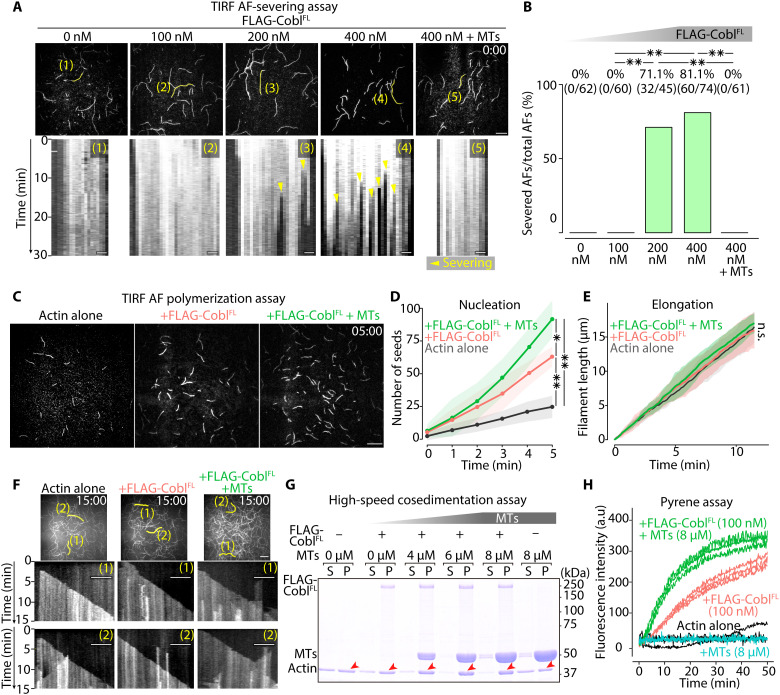
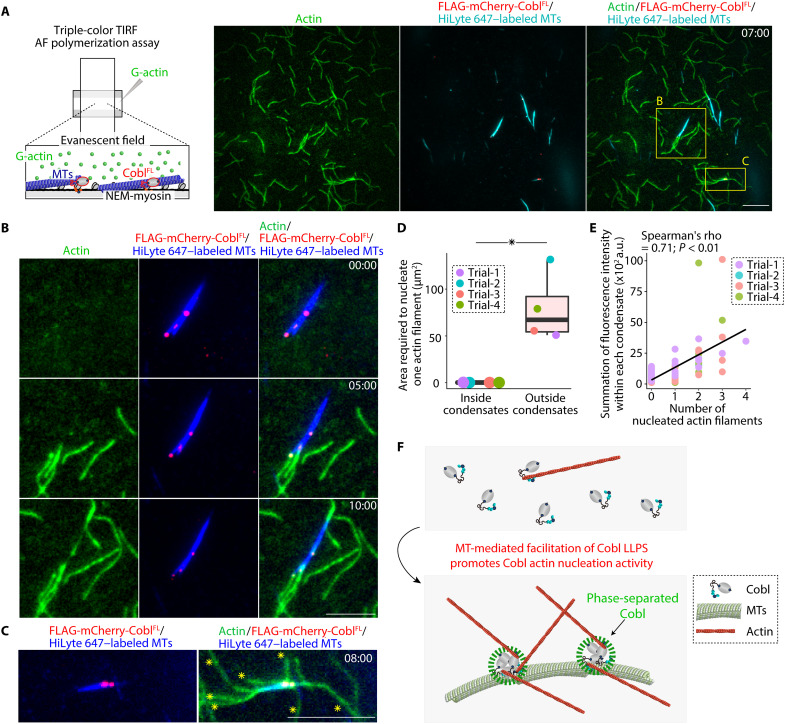
Liquid-liquid phase separation (LLPS) is involved in various dynamic biological phenomena. In epithelial cells, dynamic regulation of junctional actin filaments tethered to the apical junctional complex (AJC) is critical for maintaining internal homeostasis against external perturbations; however, the role of LLPS in this process remains unknown. Here, after identifying a multifunctional actin nucleator, cordon bleu (Cobl), as an AJC-enriched microtubule-associated protein, we conducted comprehensive in vitro and in vivo analyses. We found that apical microtubules promoted LLPS of Cobl at the AJC, and Cobl actin assembly activity increased upon LLPS. Thus, microtubules spatiotemporally regulated junctional actin assembly for epithelial morphogenesis and paracellular barriers. Collectively, these findings established that LLPS of the actin nucleator Cobl mediated dynamic microtubule-actin cross-talk in junctions, which fine-tuned the epithelial barrier.
Figures

References
-
- Y. Shin, C. P. Brangwynne, Liquid phase condensation in cell physiology and disease. Science 357, eaaf4382 (2017). - PubMed
-
- O. Beutel, R. Maraspini, K. Pombo-García, C. Martin-Lemaitre, A. Honigmann, Phase separation of zonula occludens proteins drives formation of tight junctions. Cell 179, 923–936.e11 (2019). - PubMed
-
- S. Citi, Cell biology: Tight junctions as biomolecular condensates. Curr. Biol. 30, R83–R86 (2020). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
