

A Cre-deleter specific for embryo-derived brain macrophages reveals distinct features of microglia and border macrophages
- PMID: 36791722
- PMCID: PMC10175109
- DOI: 10.1016/j.immuni.2023.01.028
A Cre-deleter specific for embryo-derived brain macrophages reveals distinct features of microglia and border macrophages
Abstract
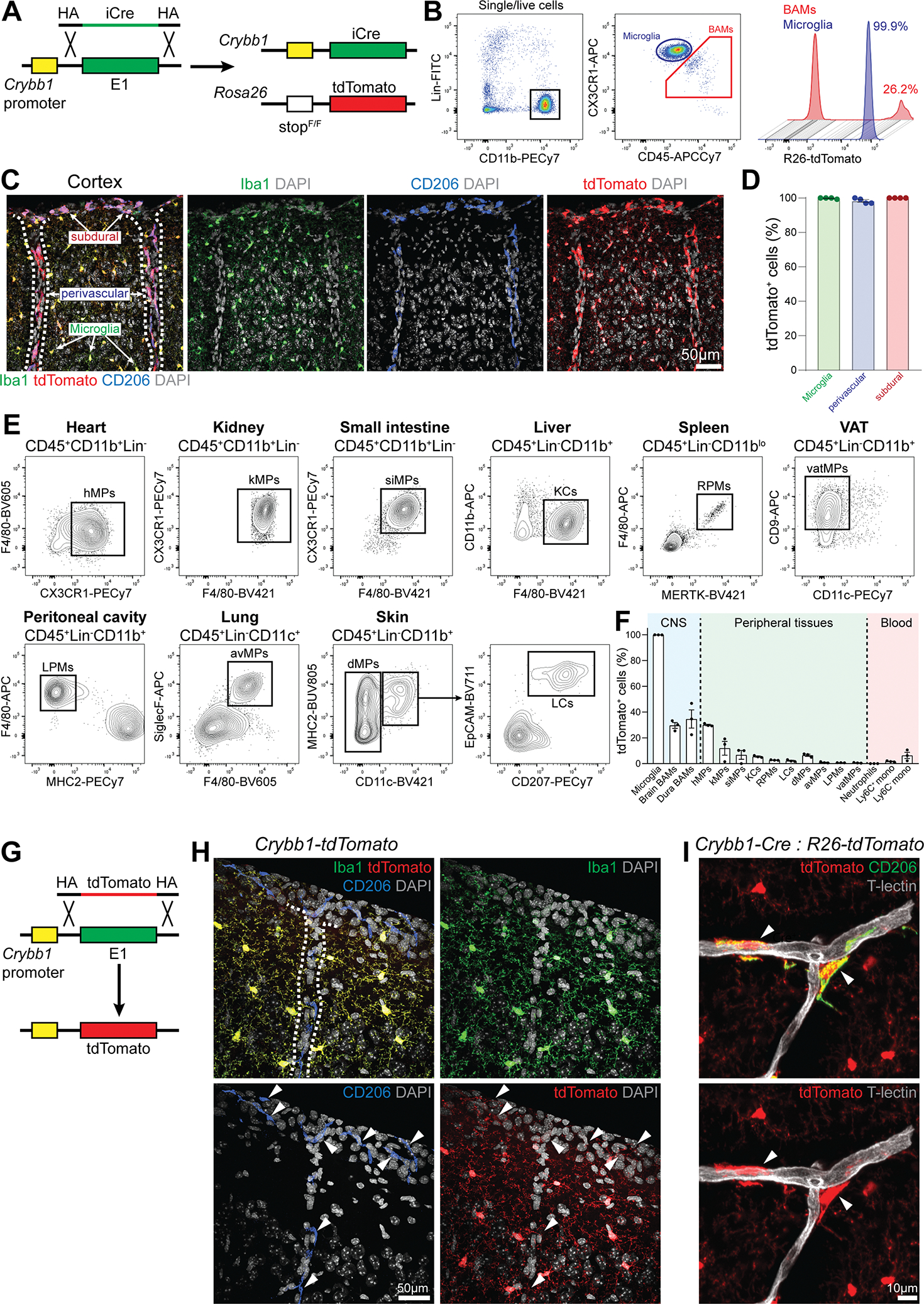
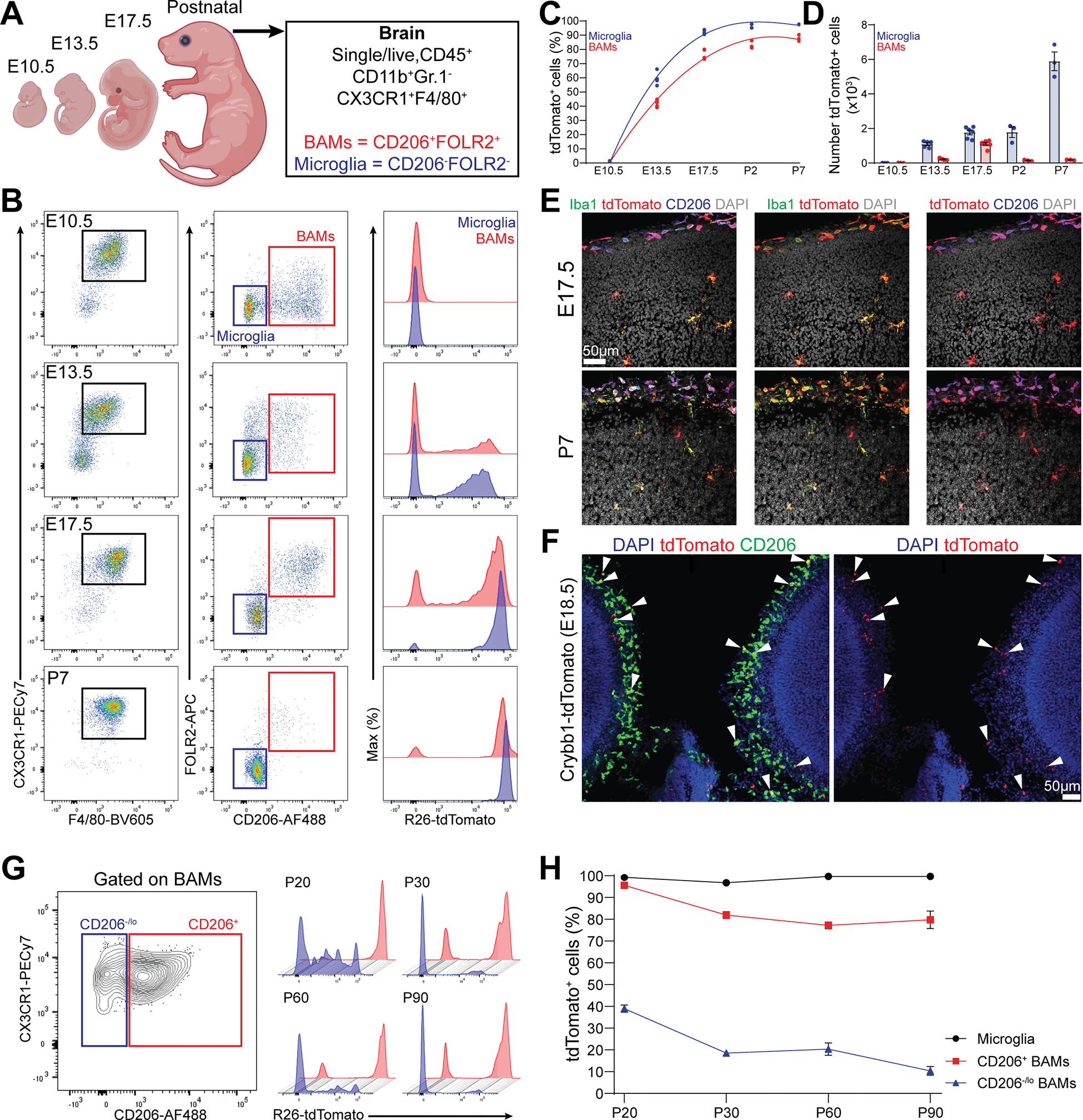
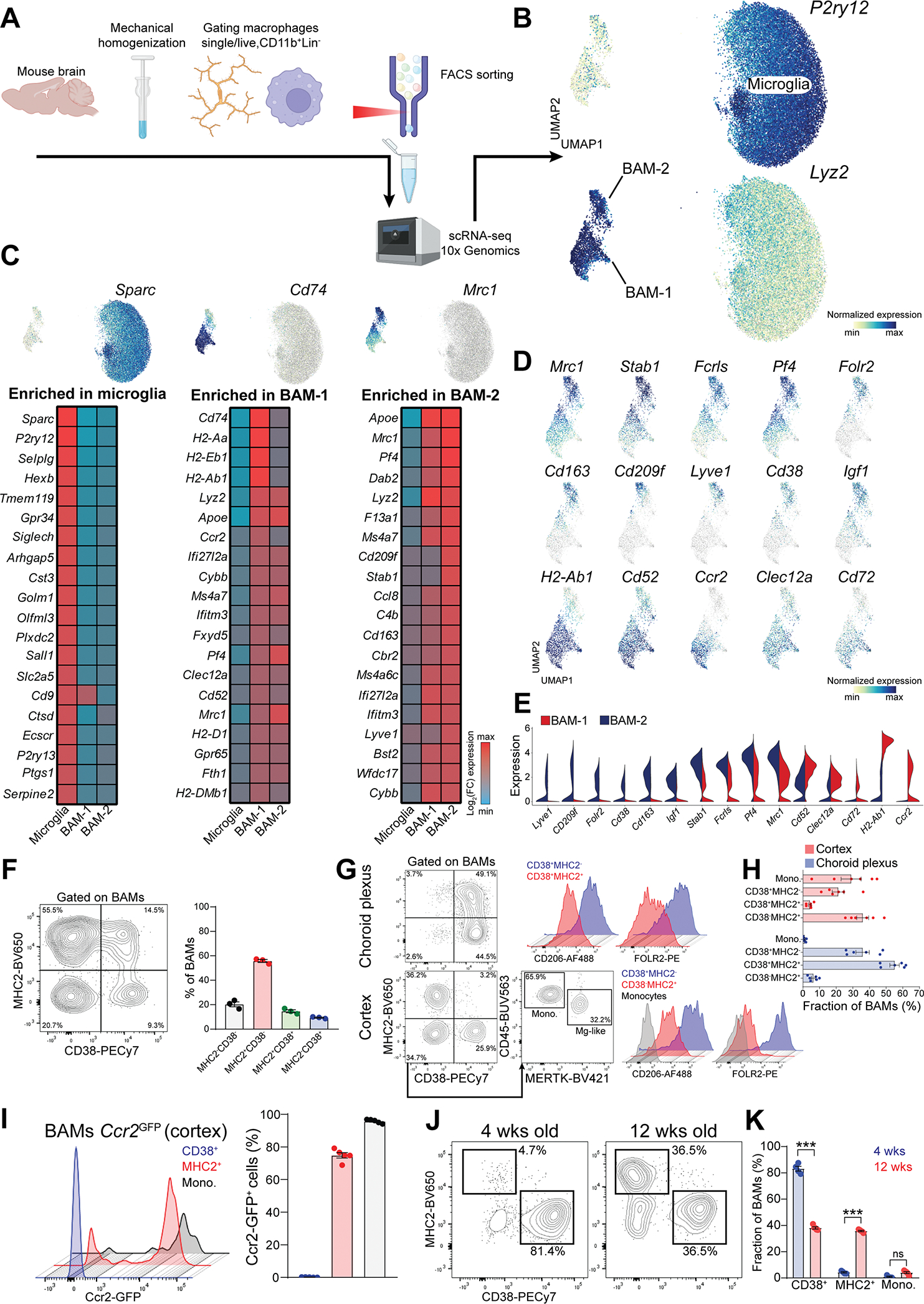
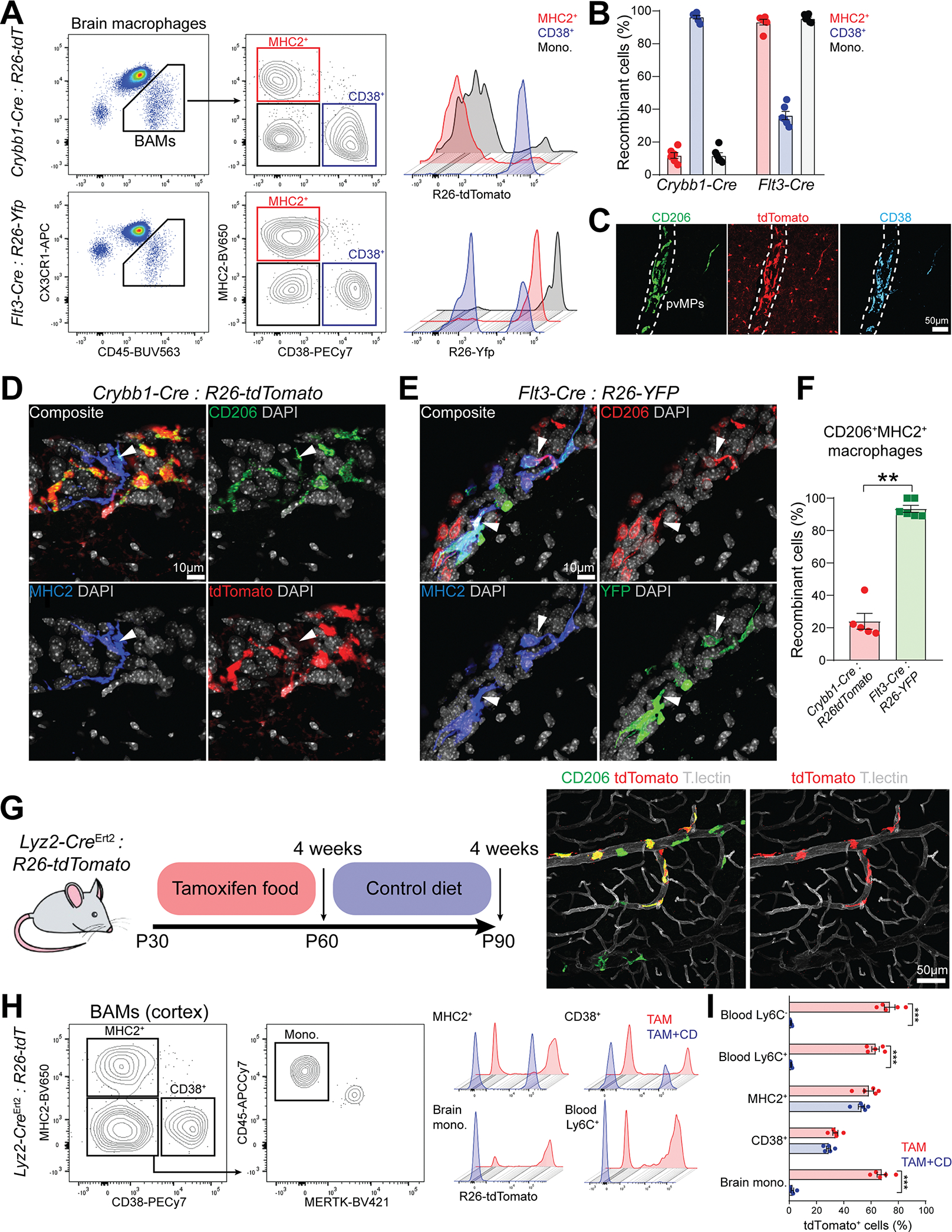
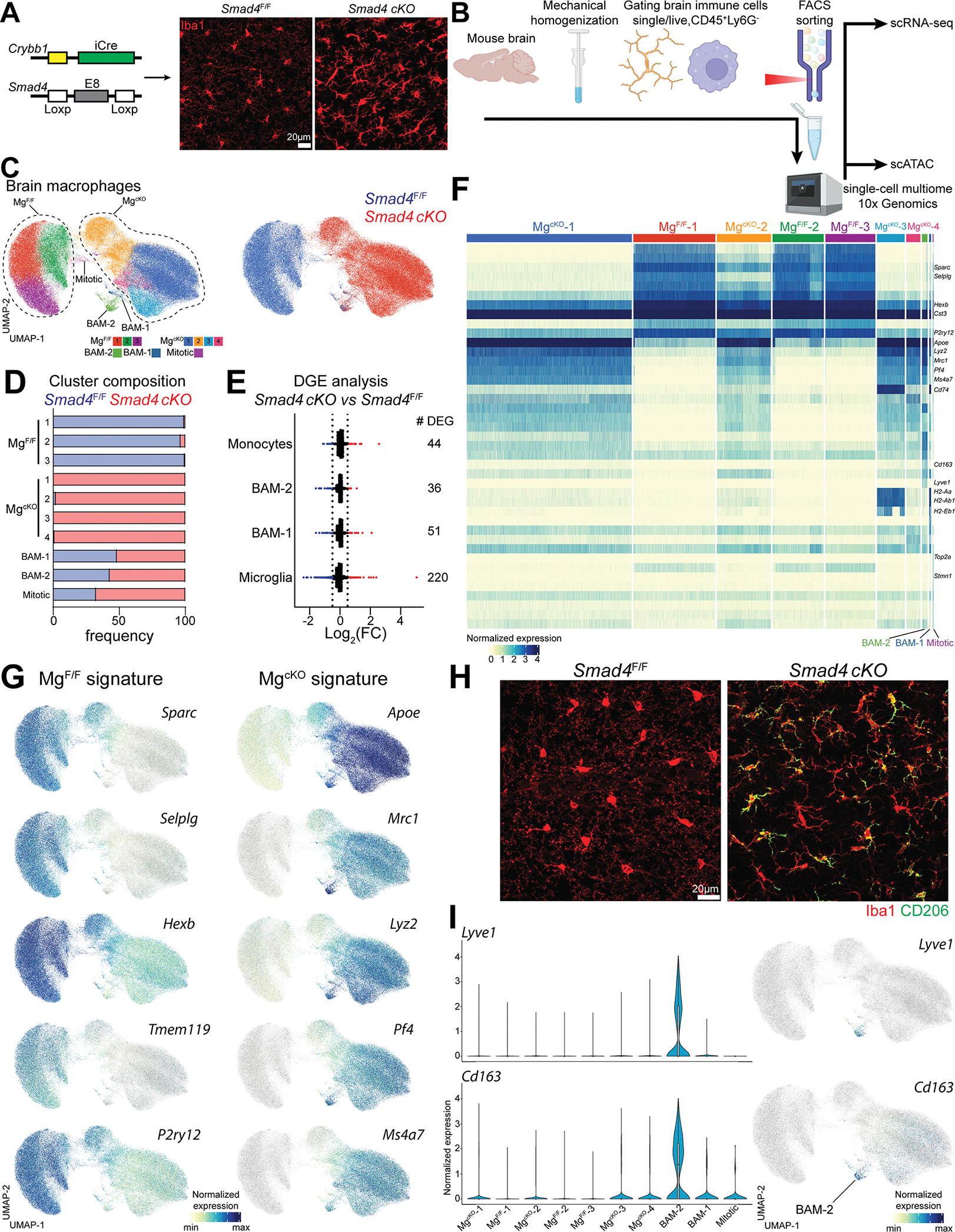
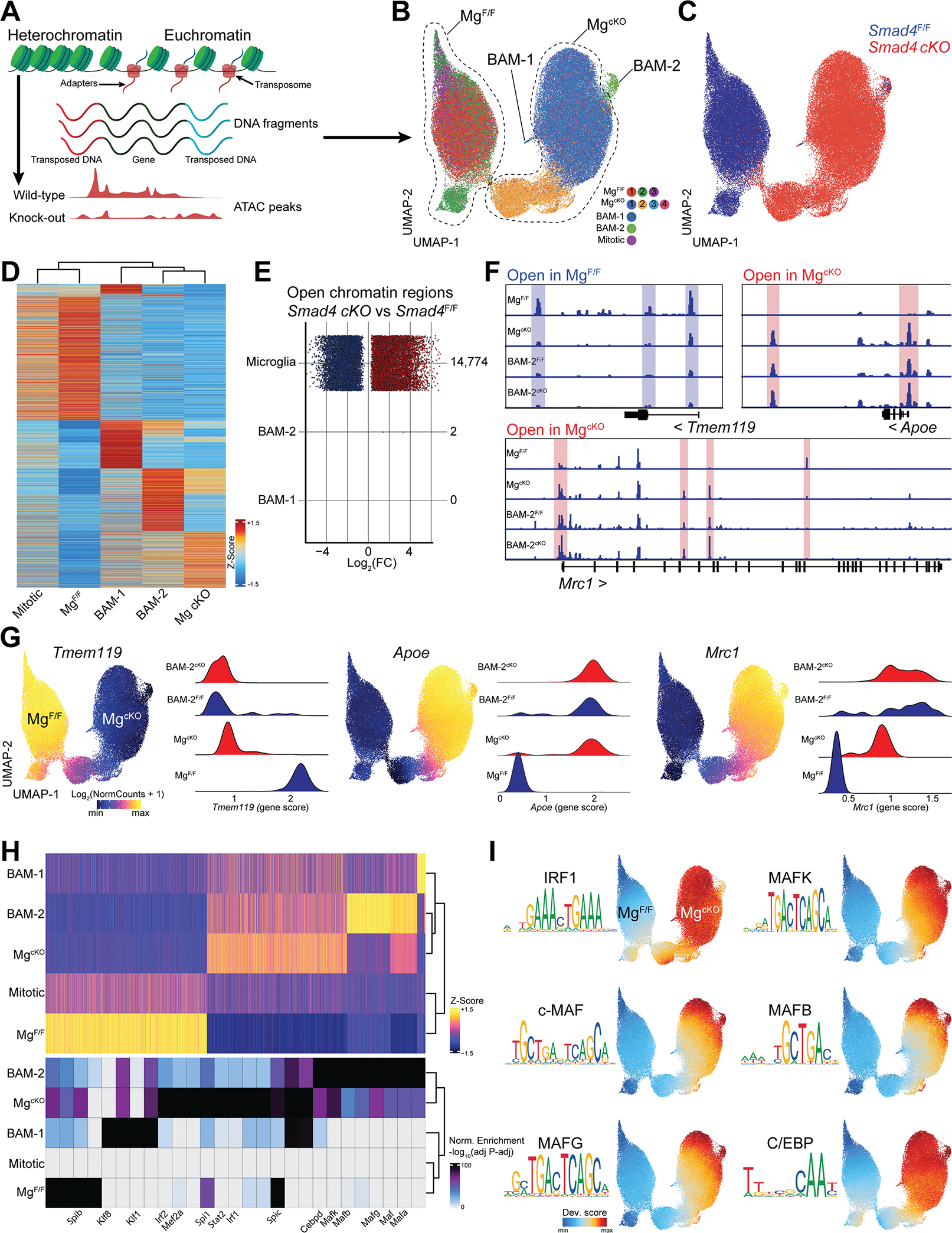
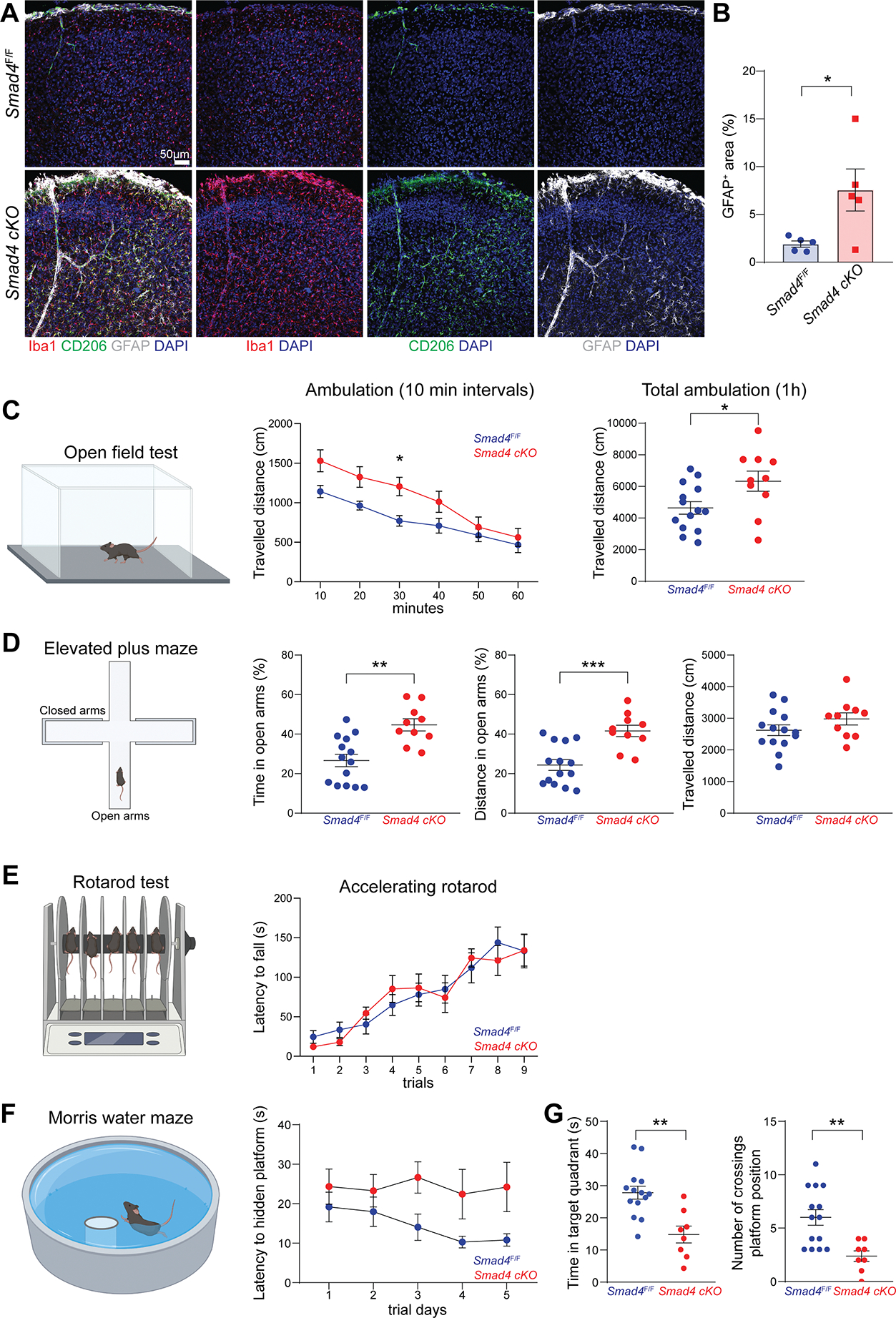
Genetic tools to target microglia specifically and efficiently from the early stages of embryonic development are lacking. We generated a constitutive Cre line controlled by the microglia signature gene Crybb1 that produced nearly complete recombination in embryonic brain macrophages (microglia and border-associated macrophages [BAMs]) by the perinatal period, with limited recombination in peripheral myeloid cells. Using this tool in combination with Flt3-Cre lineage tracer, single-cell RNA-sequencing analysis, and confocal imaging, we resolved embryonic-derived versus monocyte-derived BAMs in the mouse cortex. Deletion of the transcription factor SMAD4 in microglia and embryonic-derived BAMs using Crybb1-Cre caused a developmental arrest of microglia, which instead acquired a BAM specification signature. By contrast, the development of genuine BAMs remained unaffected. Our results reveal that SMAD4 drives a transcriptional and epigenetic program that is indispensable for the commitment of brain macrophages to the microglia fate and highlight Crybb1-Cre as a tool for targeting embryonic brain macrophages.
Keywords: BAMs; SMAD4; fate-mapping; macrophages ontogeny; microglia.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests A.T.S. is a founder of Immunai and Cartography Biosciences and receives research funding from Allogene Therapeutics and Merck Research Laboratories.
Figures
References
-
- Kierdorf K, Masuda T, Jordão MJC, Prinz M (2019) Macrophages at CNS interfaces: ontogeny and function in health and disease. Nat Rev Neurosci 20:547–562 - PubMed
-
- Schulz C, Gomez Perdiguero E, Chorro L, et al. (2012) A lineage of myeloid cells independent of Myb and hematopoietic stem cells. Science 336:86–90 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
