

Self-inhibited State of Venezuelan Equine Encephalitis Virus (VEEV) nsP2 Cysteine Protease: A Crystallographic and Molecular Dynamics Analysis
- PMID: 36792007
- PMCID: PMC10758287
- DOI: 10.1016/j.jmb.2023.168012
Self-inhibited State of Venezuelan Equine Encephalitis Virus (VEEV) nsP2 Cysteine Protease: A Crystallographic and Molecular Dynamics Analysis
Abstract
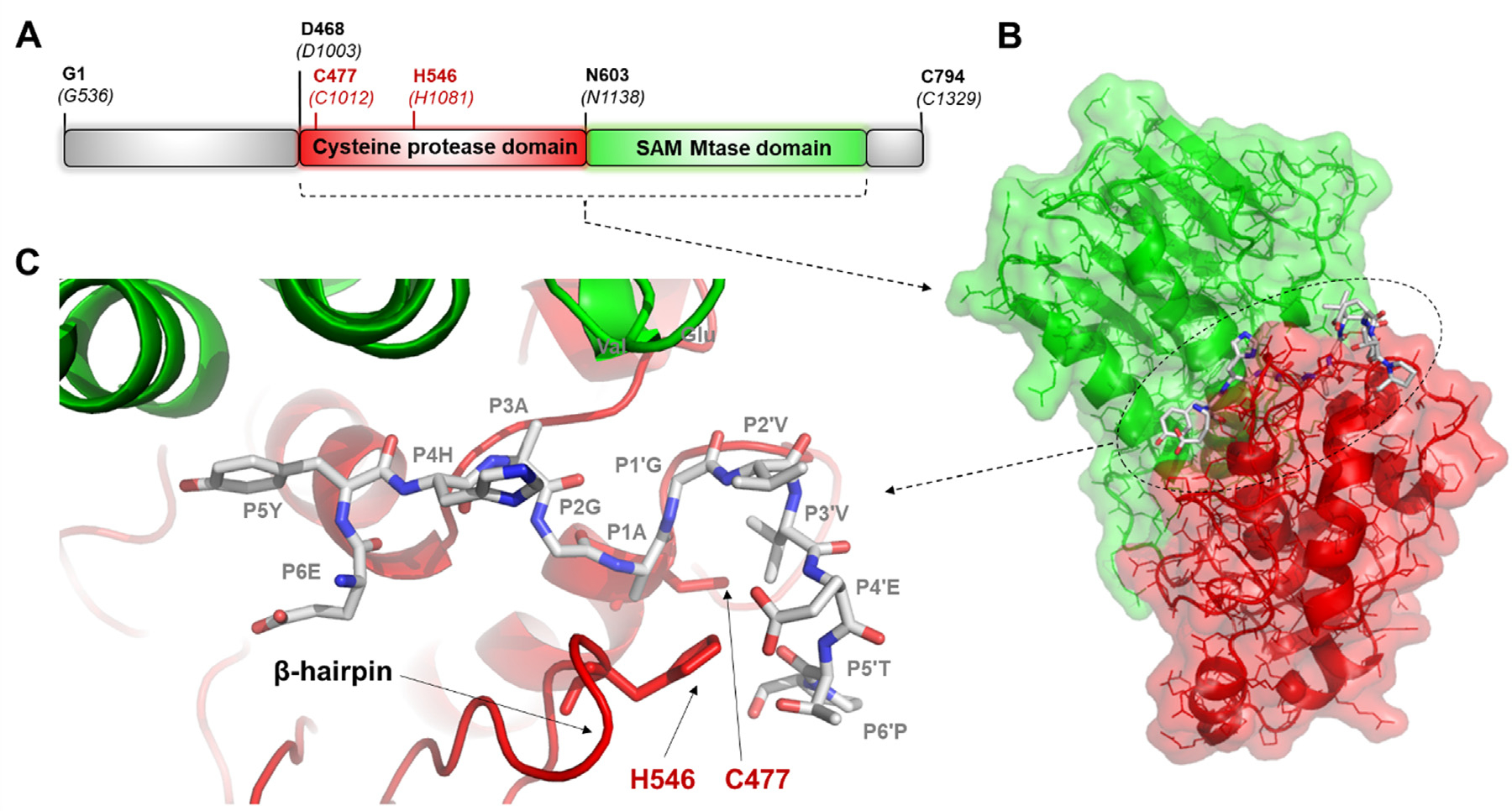
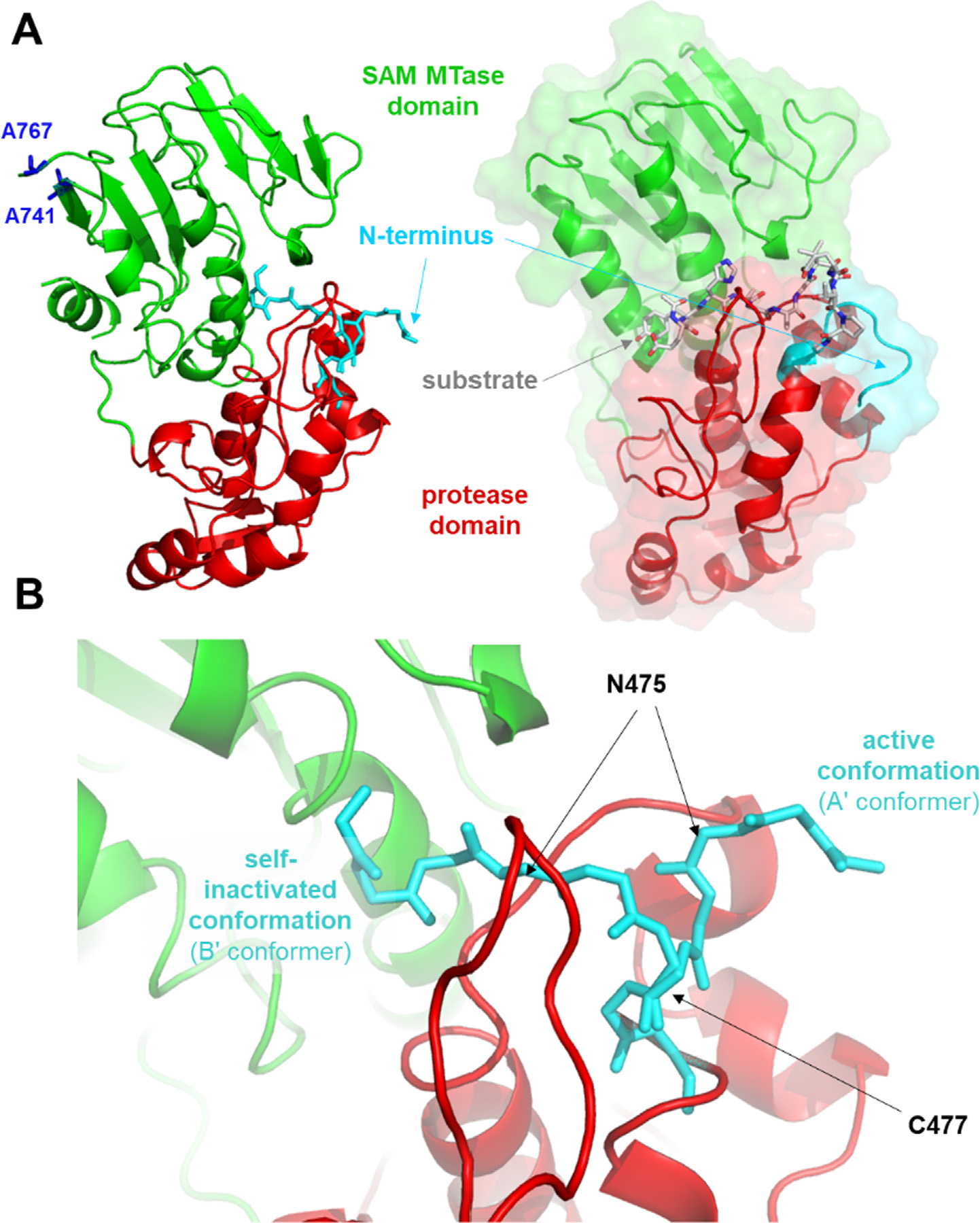
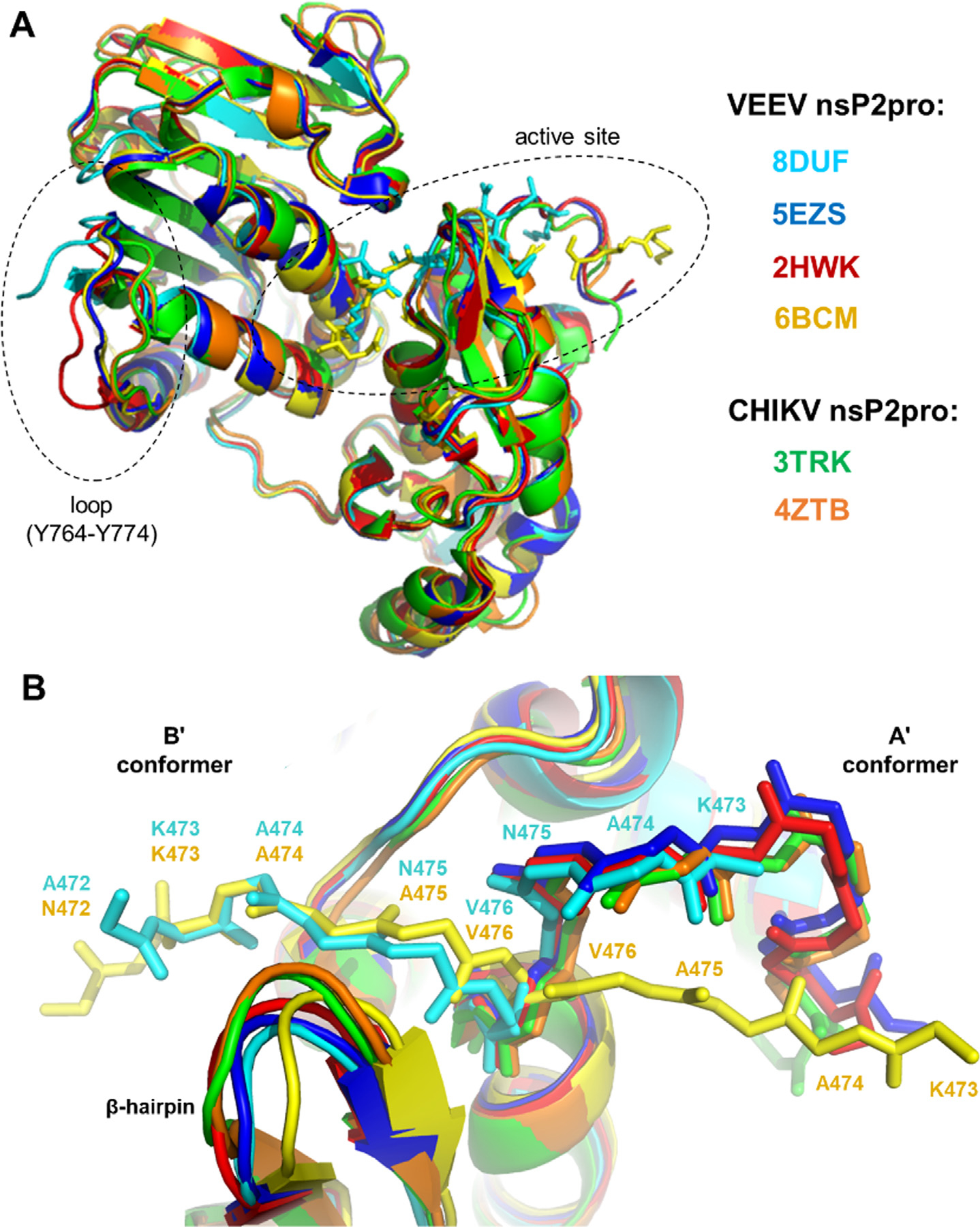
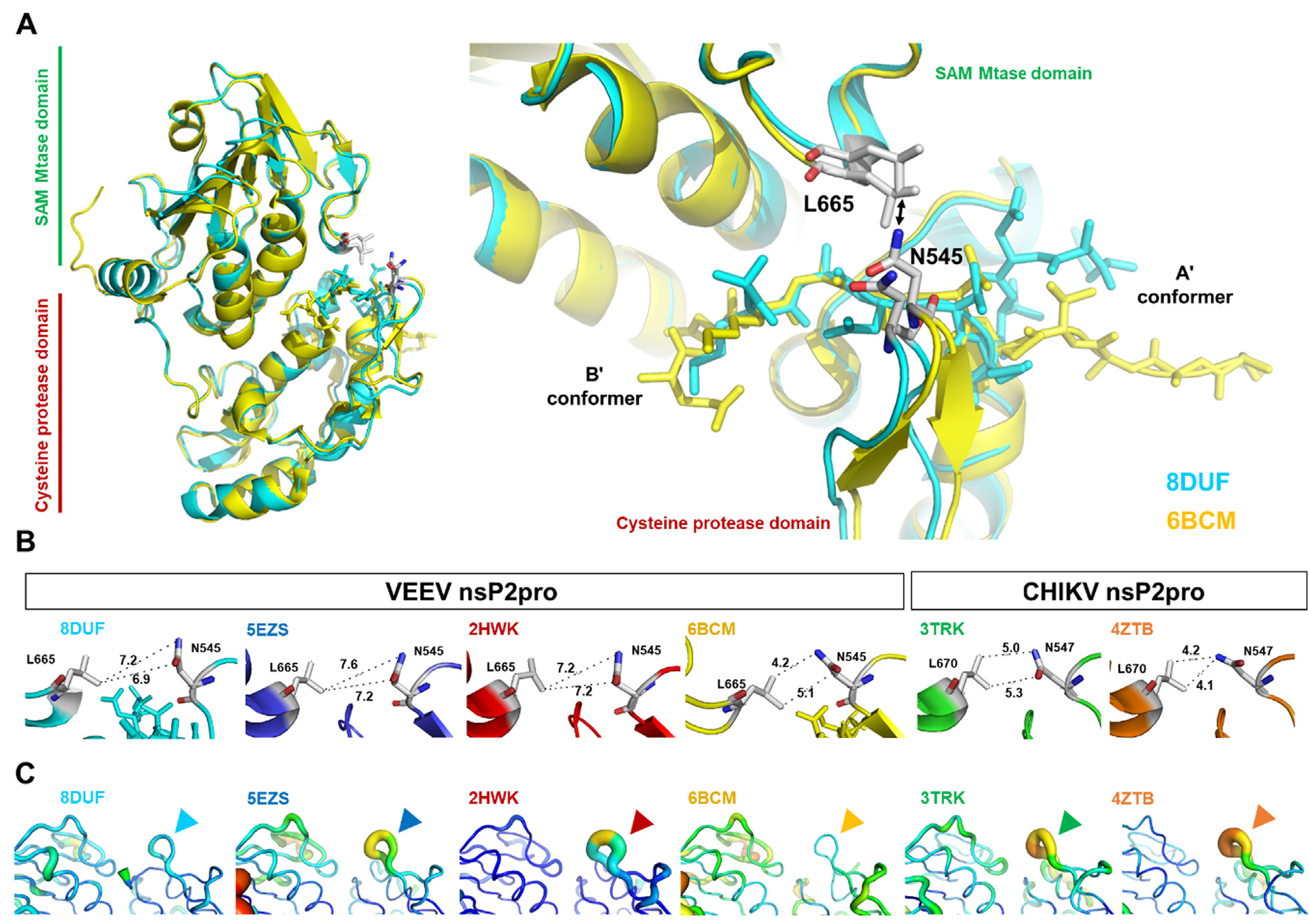
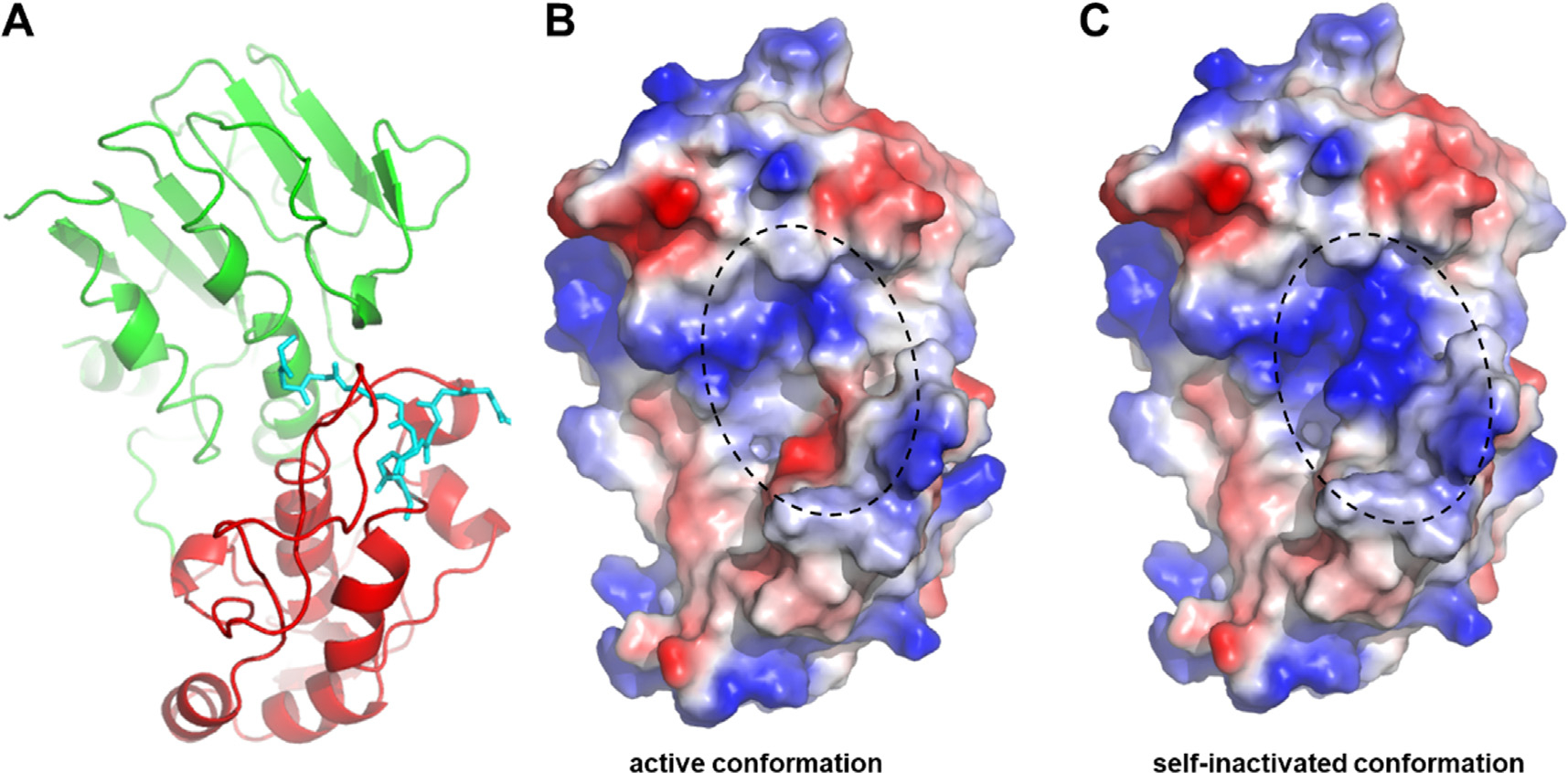
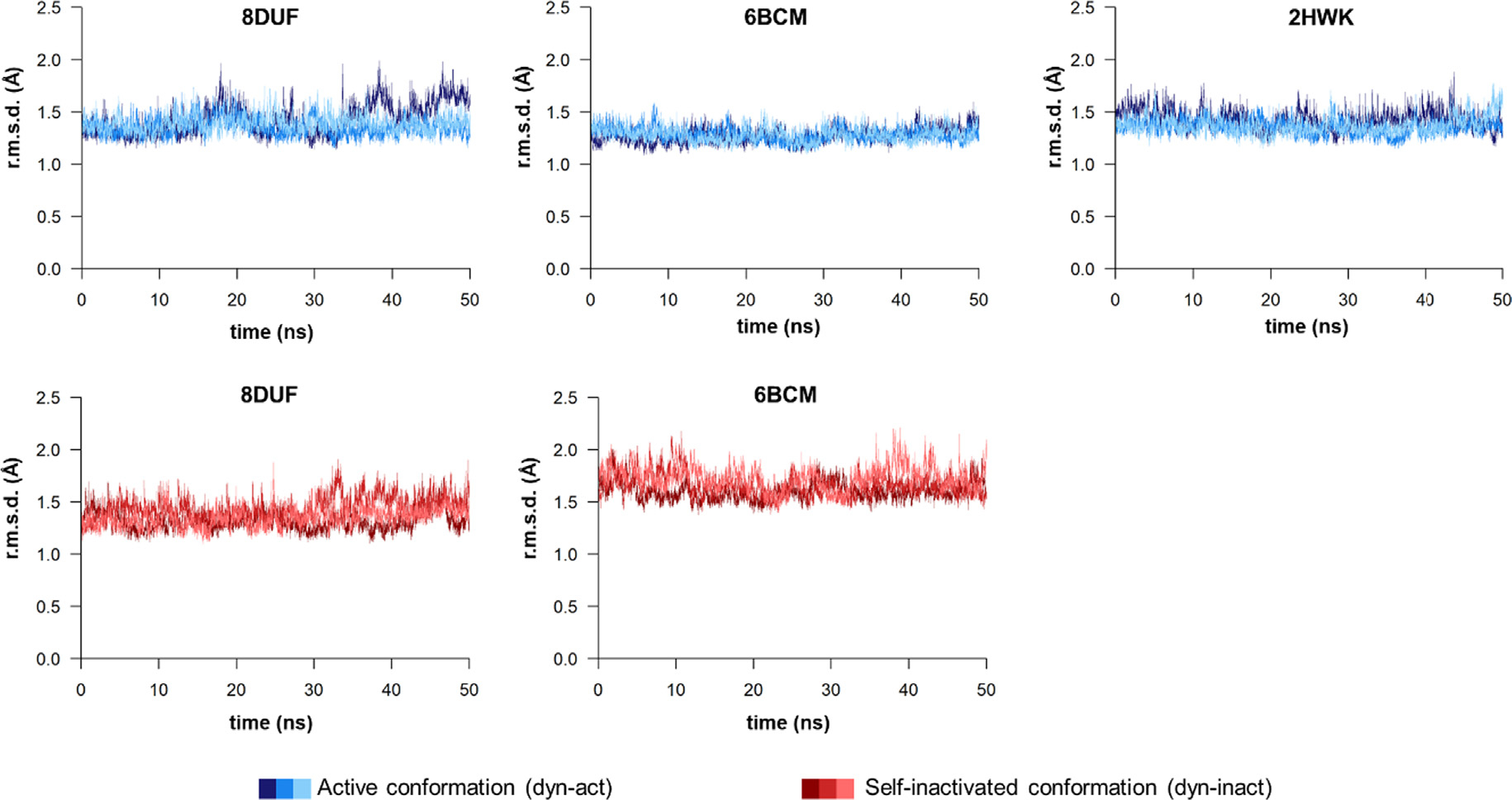
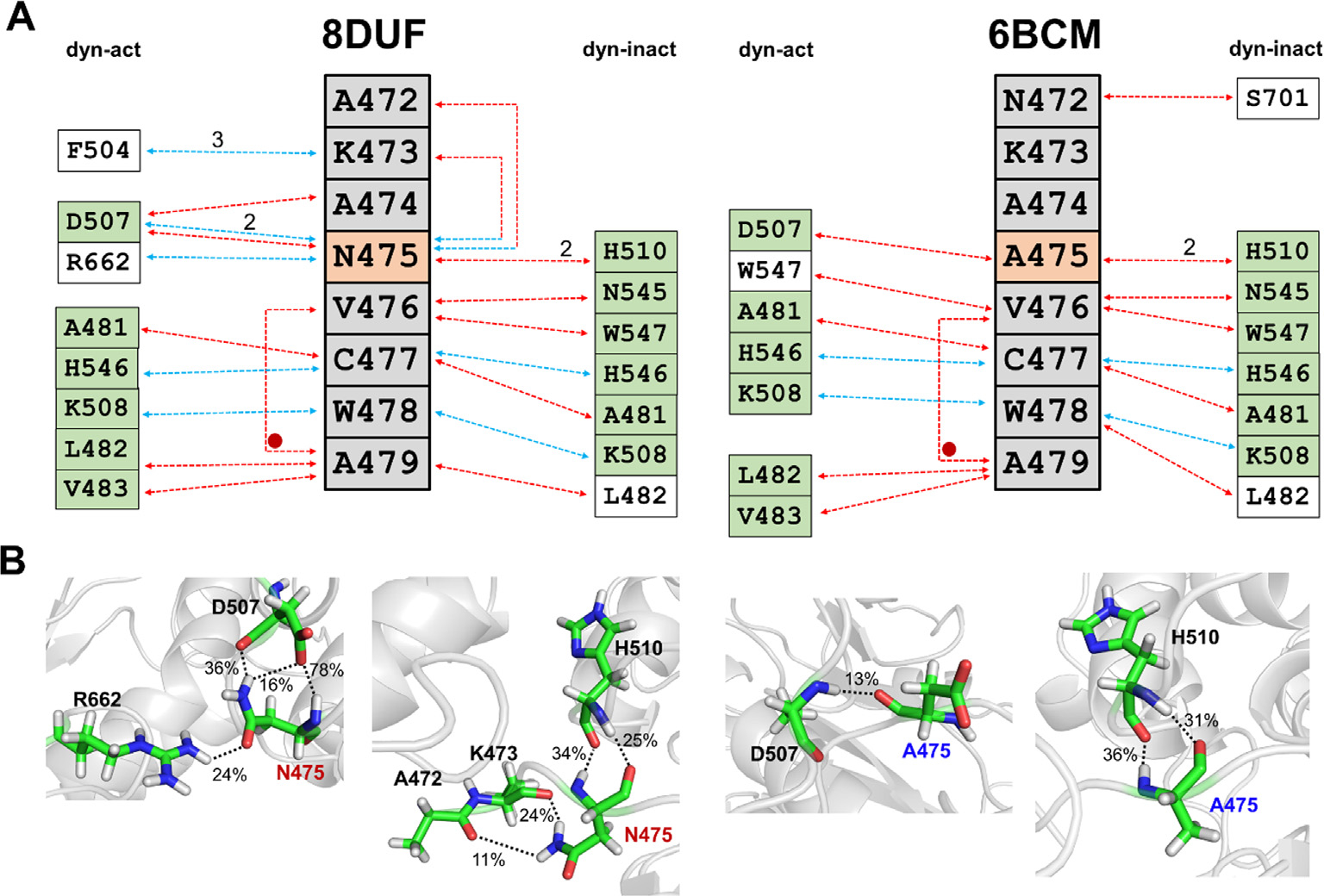
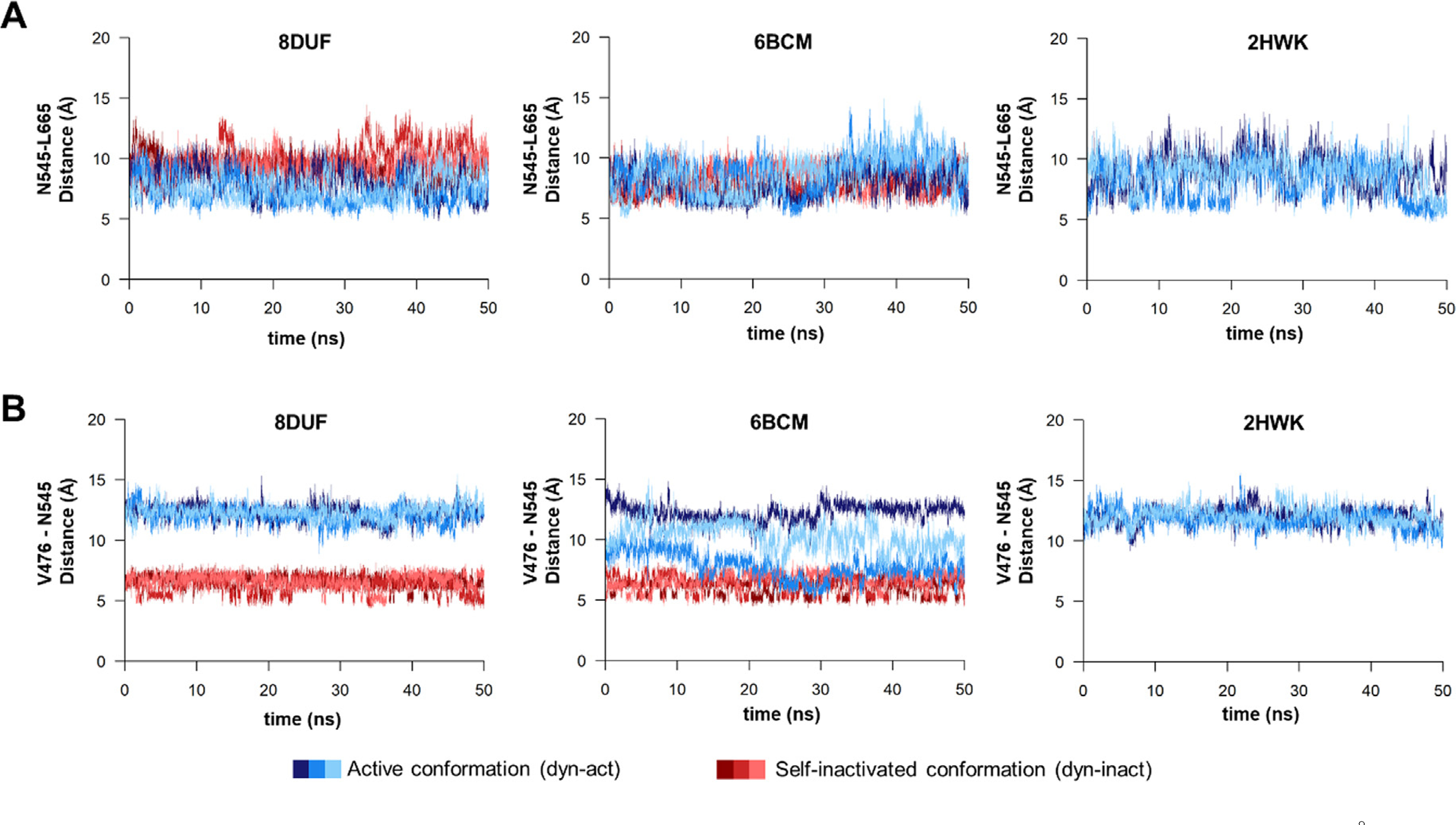
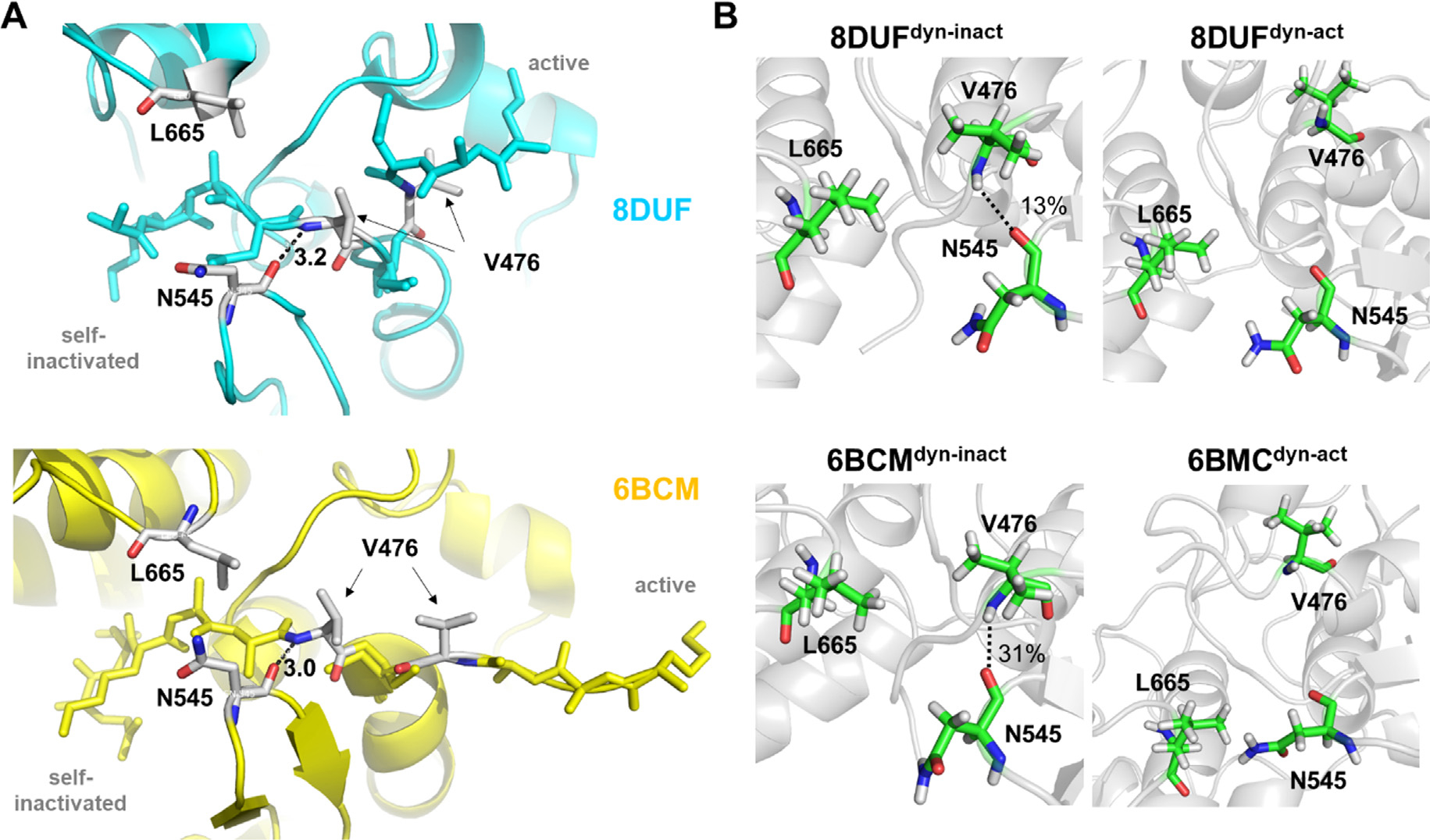
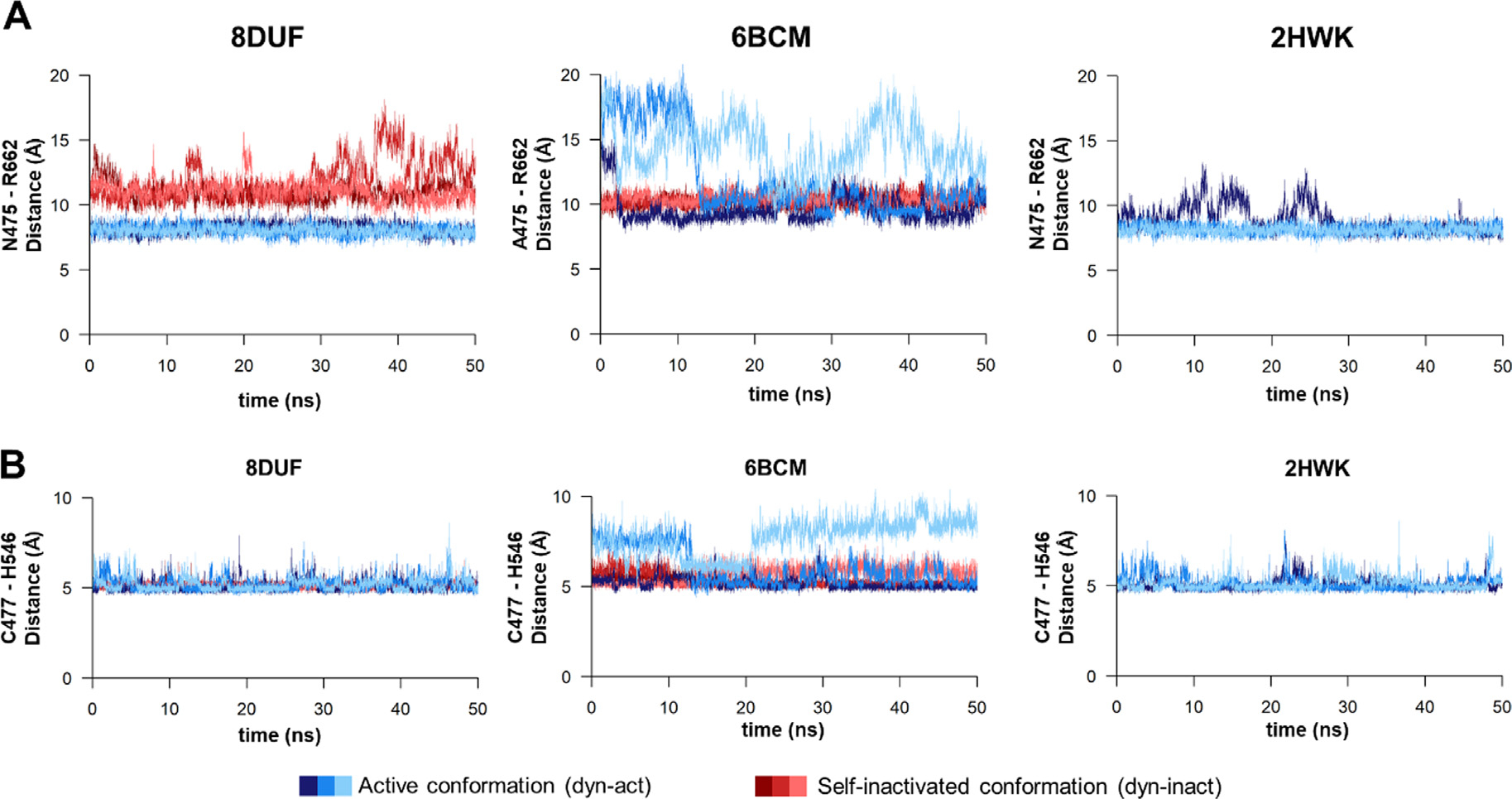
The Venezuelan equine encephalitis virus (VEEV) belongs to the Togaviridae family and is pathogenic to both humans and equines. The VEEV non-structural protein 2 (nsP2) is a cysteine protease (nsP2pro) that processes the polyprotein and thus it is a drug target for inhibitor discovery. The atomic structure of the VEEV nsP2 catalytic domain was previously characterized by both X-ray crystallography and computational studies. A modified nsP2pro harboring a N475A mutation in the N terminus was observed to exhibit an unexpected conformation: the N-terminal residues bind to the active site, mimicking binding of a substrate. The large conformational change of the N terminus was assumed to be induced by the N475A mutation, as N475 has an important role in stabilization of the N terminus and the active site. This conformation was first observed in the N475A mutant, but we also found it while determining a crystal structure of the catalytically active nsP2pro containing the wild-type N475 active site residue and K741A/K767A surface entropy reduction mutations. This suggests that the N475A mutation is not a prerequisite for self-inhibition. Here, we describe a high resolution (1.46 Å) crystal structure of a truncated nsP2pro (residues 463-785, K741A/K767A) and analyze the structure further by molecular dynamics to study the active and self-inhibited conformations of nsP2pro and its N475A mutant. A comparison of the different conformations of the N-terminal residues sheds a light on the interactions that play an important role in the stabilization of the enzyme.
Keywords: Venezuelan equine encephalitis virus; alphavirus; crystallography; molecular dynamics; protease.
Copyright © 2023 The Author(s). Published by Elsevier Ltd.. All rights reserved.
Conflict of interest statement
Declaration of Competing Interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures
Similar articles
-
Kinetic, Mutational, and Structural Studies of the Venezuelan Equine Encephalitis Virus Nonstructural Protein 2 Cysteine Protease.Biochemistry. 2016 May 31;55(21):3007-19. doi: 10.1021/acs.biochem.5b00992. Epub 2016 May 19. Biochemistry. 2016. PMID: 27030368 Free PMC article.
-
Mutation of Asn-475 in the Venezuelan Equine Encephalitis Virus nsP2 Cysteine Protease Leads to a Self-Inhibited State.Biochemistry. 2017 Nov 28;56(47):6221-6230. doi: 10.1021/acs.biochem.7b00746. Epub 2017 Nov 9. Biochemistry. 2017. PMID: 29064679
-
The crystal structure of the Venezuelan equine encephalitis alphavirus nsP2 protease.Structure. 2006 Sep;14(9):1449-58. doi: 10.1016/j.str.2006.07.010. Structure. 2006. PMID: 16962975
-
Venezuelan Equine Encephalitis Virus Capsid-The Clever Caper.Viruses. 2017 Sep 29;9(10):279. doi: 10.3390/v9100279. Viruses. 2017. PMID: 28961161 Free PMC article. Review.
-
Current Understanding of the Molecular Basis of Venezuelan Equine Encephalitis Virus Pathogenesis and Vaccine Development.Viruses. 2019 Feb 18;11(2):164. doi: 10.3390/v11020164. Viruses. 2019. PMID: 30781656 Free PMC article. Review.
Cited by
-
N-Alkyl Sulfamates as a New Class of nsP2 Cysteine Protease Inhibitors with Broad Spectrum Antialphaviral Activity.bioRxiv [Preprint]. 2025 Jul 4:2025.06.30.662352. doi: 10.1101/2025.06.30.662352. bioRxiv. 2025. PMID: 40631155 Free PMC article. Preprint.
References
-
- Bronze MS, Huycke MM, Machado LJ, Voskuhl GW, Greenfield RA, (2002). Viral Agents as Biological Weapons and Agents of Bioterrorism. Am. J. Med. Sci. 323, 316–325. - PubMed
-
- Carey BD, Bakovic A, Callahan V, Narayanan A, Kehn-Hall K, (2019). New World alphavirus protein interactomes from a therapeutic perspective. Antiviral Res. 163, 125–139. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
