

A molecular atlas reveals the tri-sectional spinning mechanism of spider dragline silk
- PMID: 36792670
- PMCID: PMC9932165
- DOI: 10.1038/s41467-023-36545-6
A molecular atlas reveals the tri-sectional spinning mechanism of spider dragline silk
Abstract
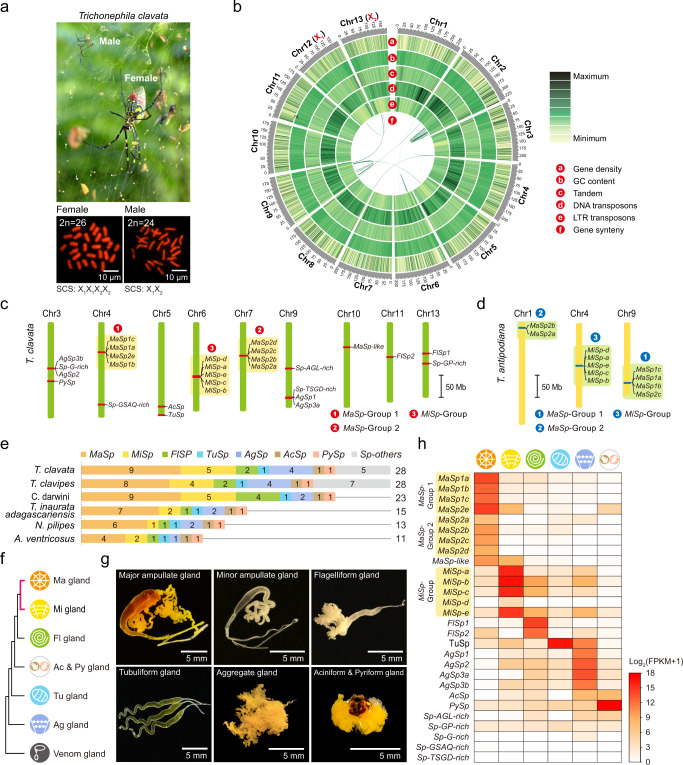
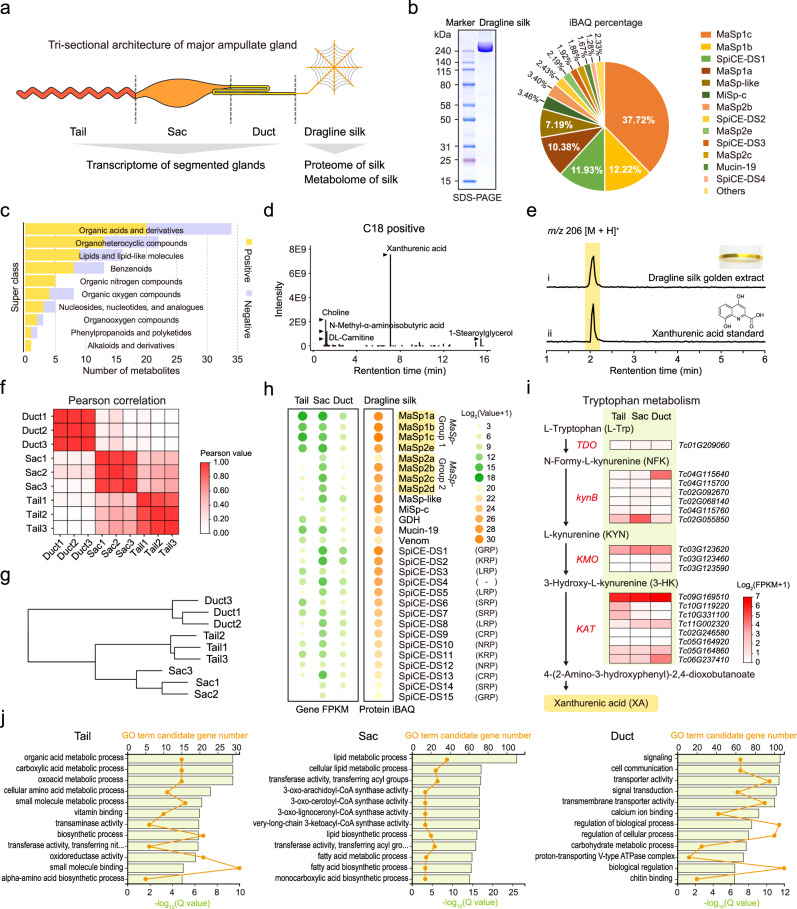
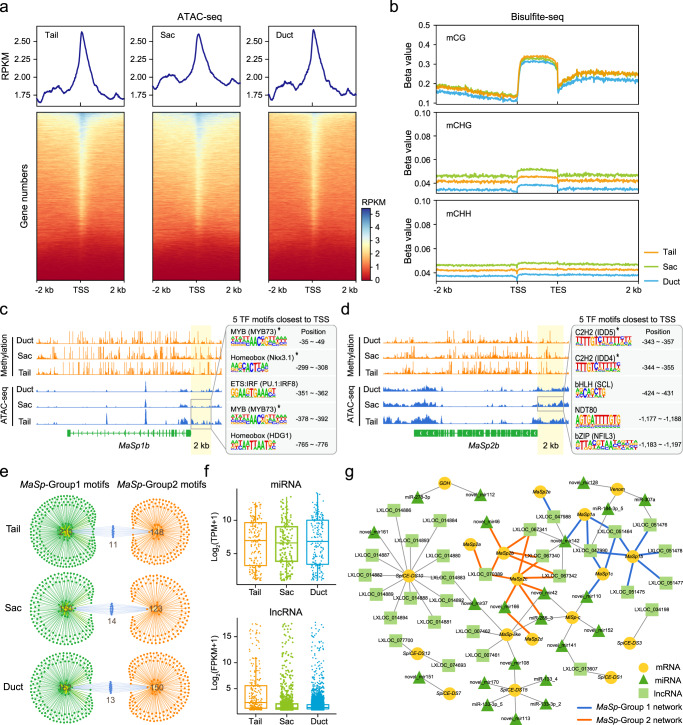
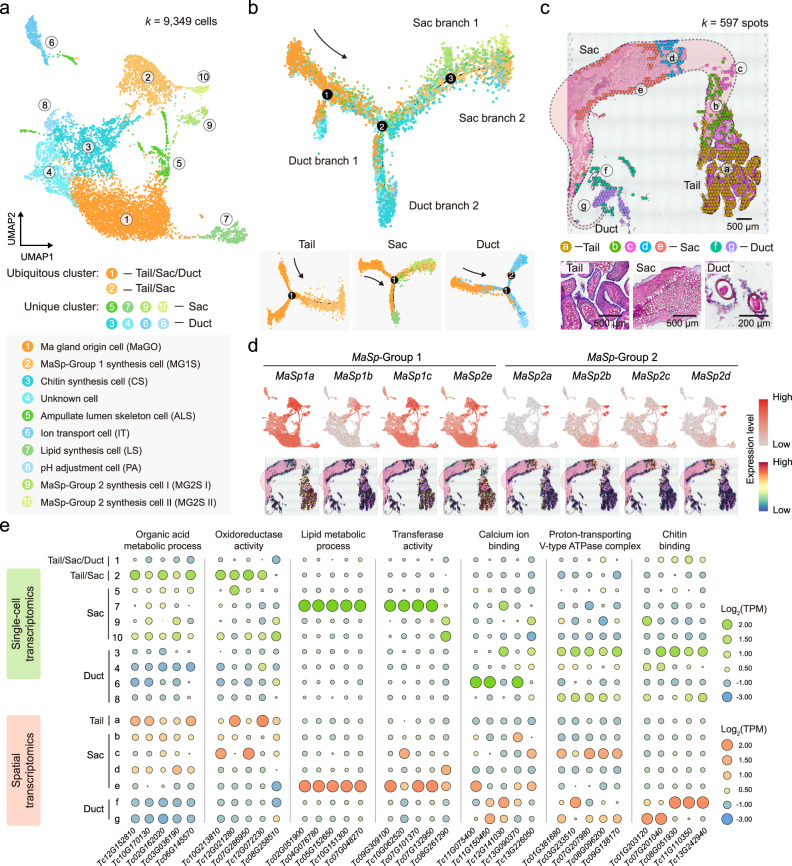
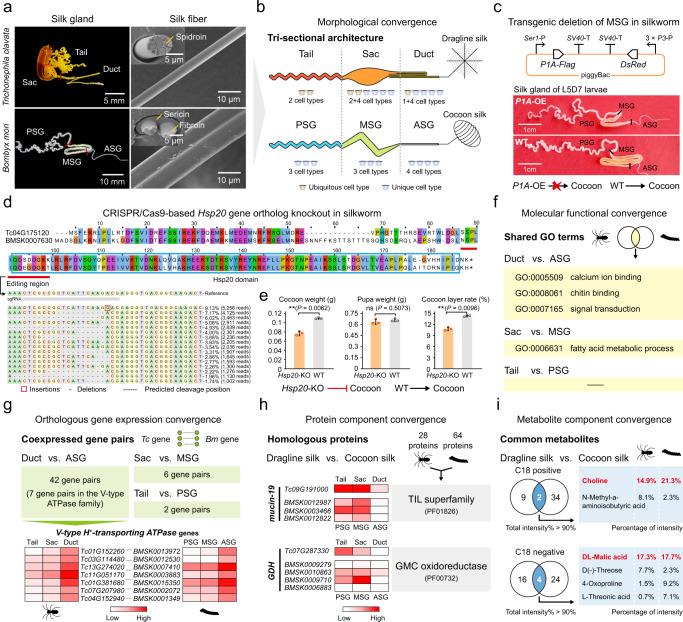
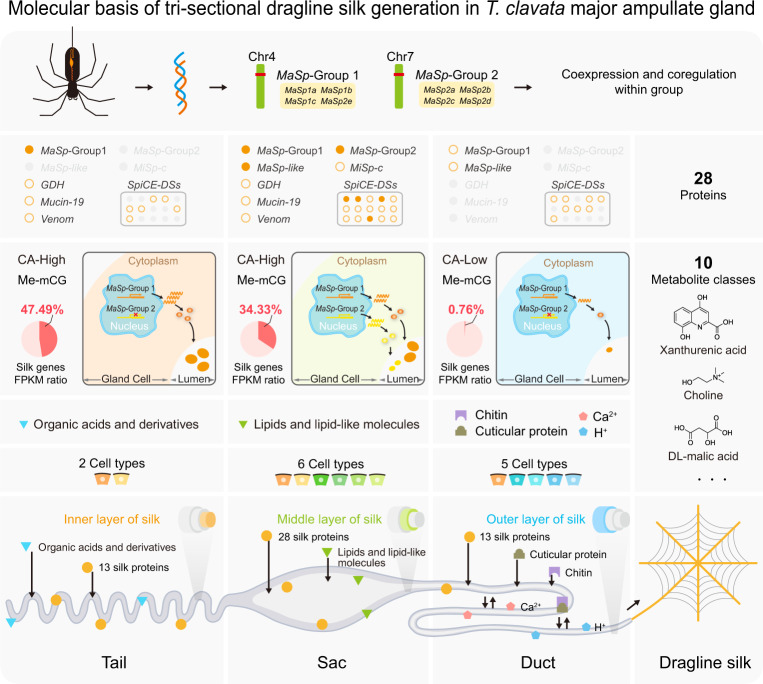
The process of natural silk production in the spider major ampullate (Ma) gland endows dragline silk with extraordinary mechanical properties and the potential for biomimetic applications. However, the precise genetic roles of the Ma gland during this process remain unknown. Here, we performed a systematic molecular atlas of dragline silk production through a high-quality genome assembly for the golden orb-weaving spider Trichonephila clavata and a multiomics approach to defining the Ma gland tri-sectional architecture: Tail, Sac, and Duct. We uncovered a hierarchical biosynthesis of spidroins, organic acids, lipids, and chitin in the sectionalized Ma gland dedicated to fine silk constitution. The ordered secretion of spidroins was achieved by the synergetic regulation of epigenetic and ceRNA signatures for genomic group-distributed spidroin genes. Single-cellular and spatial RNA profiling identified ten cell types with partitioned functional division determining the tri-sectional organization of the Ma gland. Convergence analysis and genetic manipulation further validated that this tri-sectional architecture of the silk gland was analogous across Arthropoda and inextricably linked with silk formation. Collectively, our study provides multidimensional data that significantly expand the knowledge of spider dragline silk generation and ultimately benefit innovation in spider-inspired fibers.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Multicomponent nature underlies the extraordinary mechanical properties of spider dragline silk.Proc Natl Acad Sci U S A. 2021 Aug 3;118(31):e2107065118. doi: 10.1073/pnas.2107065118. Proc Natl Acad Sci U S A. 2021. PMID: 34312234 Free PMC article.
-
Expression of spidroin proteins in the silk glands of golden orb-weaver spiders.J Exp Zool B Mol Dev Evol. 2022 Jun;338(4):241-253. doi: 10.1002/jez.b.23117. Epub 2022 Jan 4. J Exp Zool B Mol Dev Evol. 2022. PMID: 34981640
-
Classification and functional characterization of spidroin genes in a wandering spider, Pardosa pseudoannulata.Insect Biochem Mol Biol. 2022 Dec;151:103862. doi: 10.1016/j.ibmb.2022.103862. Epub 2022 Oct 31. Insect Biochem Mol Biol. 2022. PMID: 36328175
-
Structure and post-translational modifications of the web silk protein spidroin-1 from Nephila spiders.J Proteomics. 2014 Jun 13;105:174-85. doi: 10.1016/j.jprot.2014.01.002. Epub 2014 Jan 13. J Proteomics. 2014. PMID: 24434585 Review.
-
Complexity of Spider Dragline Silk.Biomacromolecules. 2022 May 9;23(5):1827-1840. doi: 10.1021/acs.biomac.1c01682. Epub 2022 Apr 4. Biomacromolecules. 2022. PMID: 35378031 Free PMC article. Review.
Cited by
-
An annotated chromosome-level genome of a troglophilic spider (Pimoa clavata).Sci Data. 2025 Aug 11;12(1):1398. doi: 10.1038/s41597-025-05703-4. Sci Data. 2025. PMID: 40790046 Free PMC article.
-
Evolution of the Spider Homeobox Gene Repertoire by Tandem and Whole Genome Duplication.Mol Biol Evol. 2023 Dec 1;40(12):msad239. doi: 10.1093/molbev/msad239. Mol Biol Evol. 2023. PMID: 37935059 Free PMC article.
-
Genomic and transcriptomic analyses of Heteropoda venatoria reveal the expansion of P450 family for starvation resistance in spiders.Gigascience. 2025 Jan 6;14:giaf019. doi: 10.1093/gigascience/giaf019. Gigascience. 2025. PMID: 40117180 Free PMC article.
-
A trade-off in evolution: the adaptive landscape of spiders without venom glands.Gigascience. 2024 Jan 2;13:giae048. doi: 10.1093/gigascience/giae048. Gigascience. 2024. PMID: 39101784 Free PMC article.
-
Origin, structure, and composition of the spider major ampullate silk fiber revealed by genomics, proteomics, and single-cell and spatial transcriptomics.Sci Adv. 2024 Aug 16;10(33):eadn0597. doi: 10.1126/sciadv.adn0597. Epub 2024 Aug 14. Sci Adv. 2024. PMID: 39141739 Free PMC article.
References
-
- Natural History Museum Bern. World Spider Catalog. Version 23.5. http://wsc.nmbe.ch (accessed 17 July 2022).
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
