

Ten years of demographic modelling of divergence and speciation in the sea
- PMID: 36793688
- PMCID: PMC9923478
- DOI: 10.1111/eva.13428
Ten years of demographic modelling of divergence and speciation in the sea
Abstract
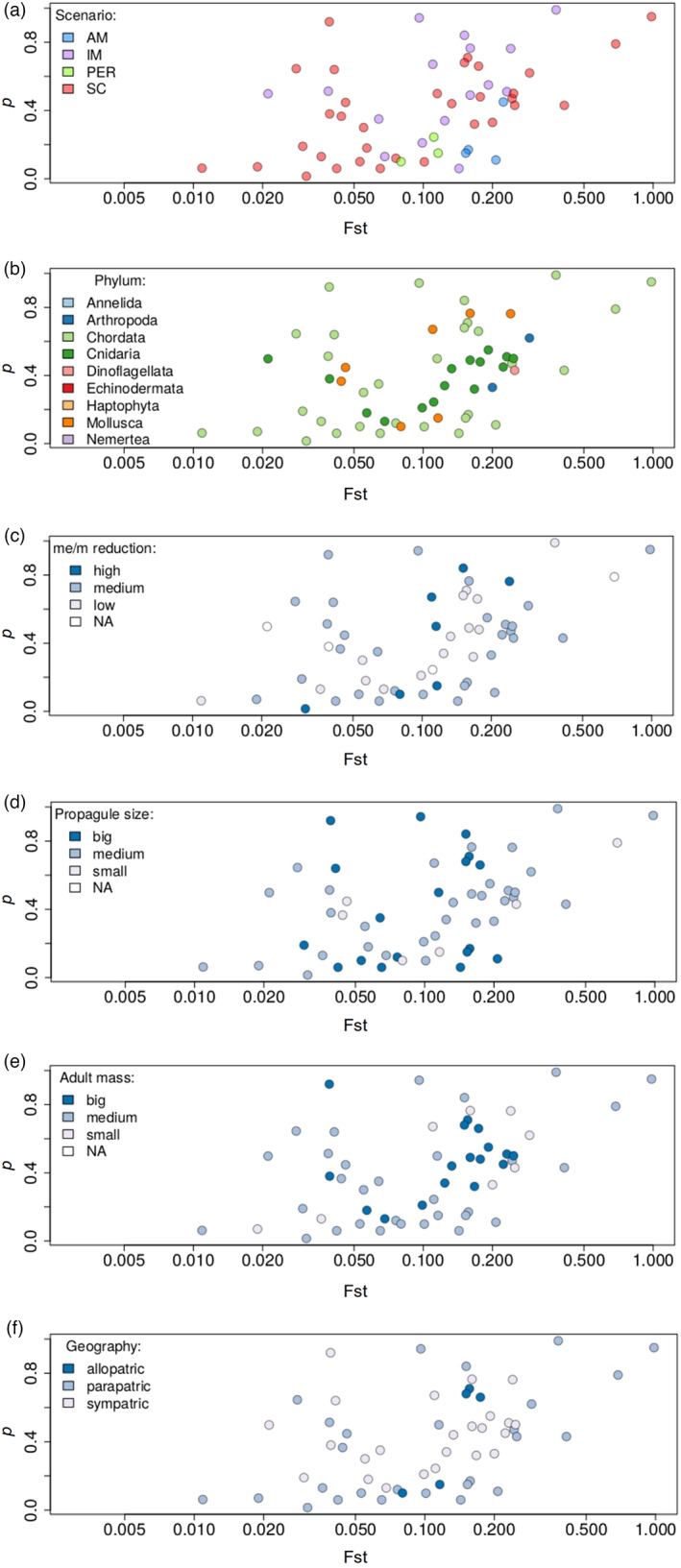
Understanding population divergence that eventually leads to speciation is essential for evolutionary biology. High species diversity in the sea was regarded as a paradox when strict allopatry was considered necessary for most speciation events because geographical barriers seemed largely absent in the sea, and many marine species have high dispersal capacities. Combining genome-wide data with demographic modelling to infer the demographic history of divergence has introduced new ways to address this classical issue. These models assume an ancestral population that splits into two subpopulations diverging according to different scenarios that allow tests for periods of gene flow. Models can also test for heterogeneities in population sizes and migration rates along the genome to account, respectively, for background selection and selection against introgressed ancestry. To investigate how barriers to gene flow arise in the sea, we compiled studies modelling the demographic history of divergence in marine organisms and extracted preferred demographic scenarios together with estimates of demographic parameters. These studies show that geographical barriers to gene flow do exist in the sea but that divergence can also occur without strict isolation. Heterogeneity of gene flow was detected in most population pairs suggesting the predominance of semipermeable barriers during divergence. We found a weak positive relationship between the fraction of the genome experiencing reduced gene flow and levels of genome-wide differentiation. Furthermore, we found that the upper bound of the 'grey zone of speciation' for our dataset extended beyond that found before, implying that gene flow between diverging taxa is possible at higher levels of divergence than previously thought. Finally, we list recommendations for further strengthening the use of demographic modelling in speciation research. These include a more balanced representation of taxa, more consistent and comprehensive modelling, clear reporting of results and simulation studies to rule out nonbiological explanations for general results.
Keywords: demographic inference; grey zone of speciation; heterogeneous gene flow; marine speciation; primary and secondary contact; reproductive isolation.
© 2022 The Authors. Evolutionary Applications published by John Wiley & Sons Ltd.
Conflict of interest statement
We have no conflict of interest to declare.
Figures





References
-
- Abbott, R. , Albach, D. , Ansell, S. , Arntzen, J. W. , Baird, S. J. E. , Bierne, N. , Boughman, J. , Brelsford, A. , Buerkle, C. A. , Buggs, R. , Butlin, R. K. , Dieckmann, U. , Eroukhmanoff, F. , Grill, A. , Cahan, S. H. , Hermansen, J. S. , Hewitt, G. , Hudson, A. G. , Jiggins, C. , … Zinner, D. (2013). Hybridization and speciation. Journal of Evolutionary Biology, 26(2), 229–246. 10.1111/j.1420-9101.2012.02599.x - DOI - PubMed
-
- Barton, N. H. (1983). Multilocus clines. Evolution, 37, 454–471. - PubMed
LinkOut - more resources
Full Text Sources
