

Reprogramming of adult human dermal fibroblasts to induced dorsal forebrain precursor cells maintains aging signatures
- PMID: 36794263
- PMCID: PMC9922835
- DOI: 10.3389/fncel.2023.1003188
Reprogramming of adult human dermal fibroblasts to induced dorsal forebrain precursor cells maintains aging signatures
Abstract
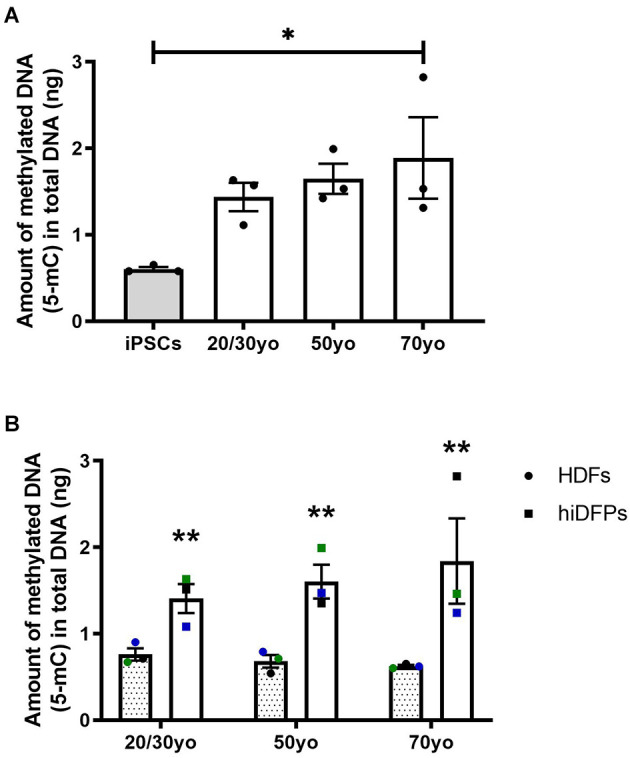
Introduction: With the increase in aging populations around the world, the development of in vitro human cell models to study neurodegenerative disease is crucial. A major limitation in using induced pluripotent stem cell (hiPSC) technology to model diseases of aging is that reprogramming fibroblasts to a pluripotent stem cell state erases age-associated features. The resulting cells show behaviors of an embryonic stage exhibiting longer telomeres, reduced oxidative stress, and mitochondrial rejuvenation, as well as epigenetic modifications, loss of abnormal nuclear morphologies, and age-associated features. Methods: We have developed a protocol utilizing stable, non-immunogenic chemically modified mRNA (cmRNA) to convert adult human dermal fibroblasts (HDFs) to human induced dorsal forebrain precursor (hiDFP) cells, which can subsequently be differentiated into cortical neurons. Analyzing an array of aging biomarkers, we demonstrate for the first time the effect of direct-to-hiDFP reprogramming on cellular age. Results: We confirm direct-to-hiDFP reprogramming does not affect telomere length or the expression of key aging markers. However, while direct-to-hiDFP reprogramming does not affect senescence-associated β-galactosidase activity, it enhances the level of mitochondrial reactive oxygen species and the amount of DNA methylation compared to HDFs. Interestingly, following neuronal differentiation of hiDFPs we observed an increase in cell soma size as well as neurite number, length, and branching with increasing donor age suggesting that neuronal morphology is altered with age. Discussion: We propose direct-to-hiDFP reprogramming provides a strategy for modeling age-associated neurodegenerative diseases allowing the persistence of age-associated signatures not seen in hiPSC-derived cultures, thereby facilitating our understanding of neurodegenerative disease and identification of therapeutic targets.
Keywords: DNA methylation; direct cell reprogramming; human induced dorsal forebrain precursors; induced pluripotent stem cell; neurodegenerative disease; oxidative stress; senescence; telomere length.
Copyright © 2023 McCaughey-Chapman, Tarczyluk-Wells, Combrinck, Edwards, Jones and Connor.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures





Similar articles
-
Directly reprogrammed fragile X syndrome dorsal forebrain precursor cells generate cortical neurons exhibiting impaired neuronal maturation.Front Cell Neurosci. 2023 Sep 21;17:1254412. doi: 10.3389/fncel.2023.1254412. eCollection 2023. Front Cell Neurosci. 2023. PMID: 37810261 Free PMC article.
-
Small Molecules Enhance Reprogramming of Adult Human Dermal Fibroblasts to Dorsal Forebrain Precursor Cells.Stem Cells Dev. 2022 Feb;31(3-4):78-89. doi: 10.1089/scd.2021.0130. Epub 2022 Jan 21. Stem Cells Dev. 2022. PMID: 34963331
-
Reversibility of cellular aging by reprogramming through an embryonic-like state: a new paradigm for human cell rejuvenation.Cent Asian J Glob Health. 2014 Jan 24;2(Suppl):88. doi: 10.5195/cajgh.2013.88. eCollection 2013. Cent Asian J Glob Health. 2014. PMID: 29805851 Free PMC article.
-
Induced pluripotent stem cell-derived and directly reprogrammed neurons to study neurodegenerative diseases: The impact of aging signatures.Front Aging Neurosci. 2022 Dec 20;14:1069482. doi: 10.3389/fnagi.2022.1069482. eCollection 2022. Front Aging Neurosci. 2022. PMID: 36620769 Free PMC article. Review.
-
The aging signature: a hallmark of induced pluripotent stem cells?Aging Cell. 2014 Feb;13(1):2-7. doi: 10.1111/acel.12182. Epub 2013 Nov 21. Aging Cell. 2014. PMID: 24256351 Free PMC article. Review.
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials