

Protease allergens as initiators-regulators of allergic inflammation
- PMID: 36794967
- PMCID: PMC10159943
- DOI: 10.1111/all.15678
Protease allergens as initiators-regulators of allergic inflammation
Abstract
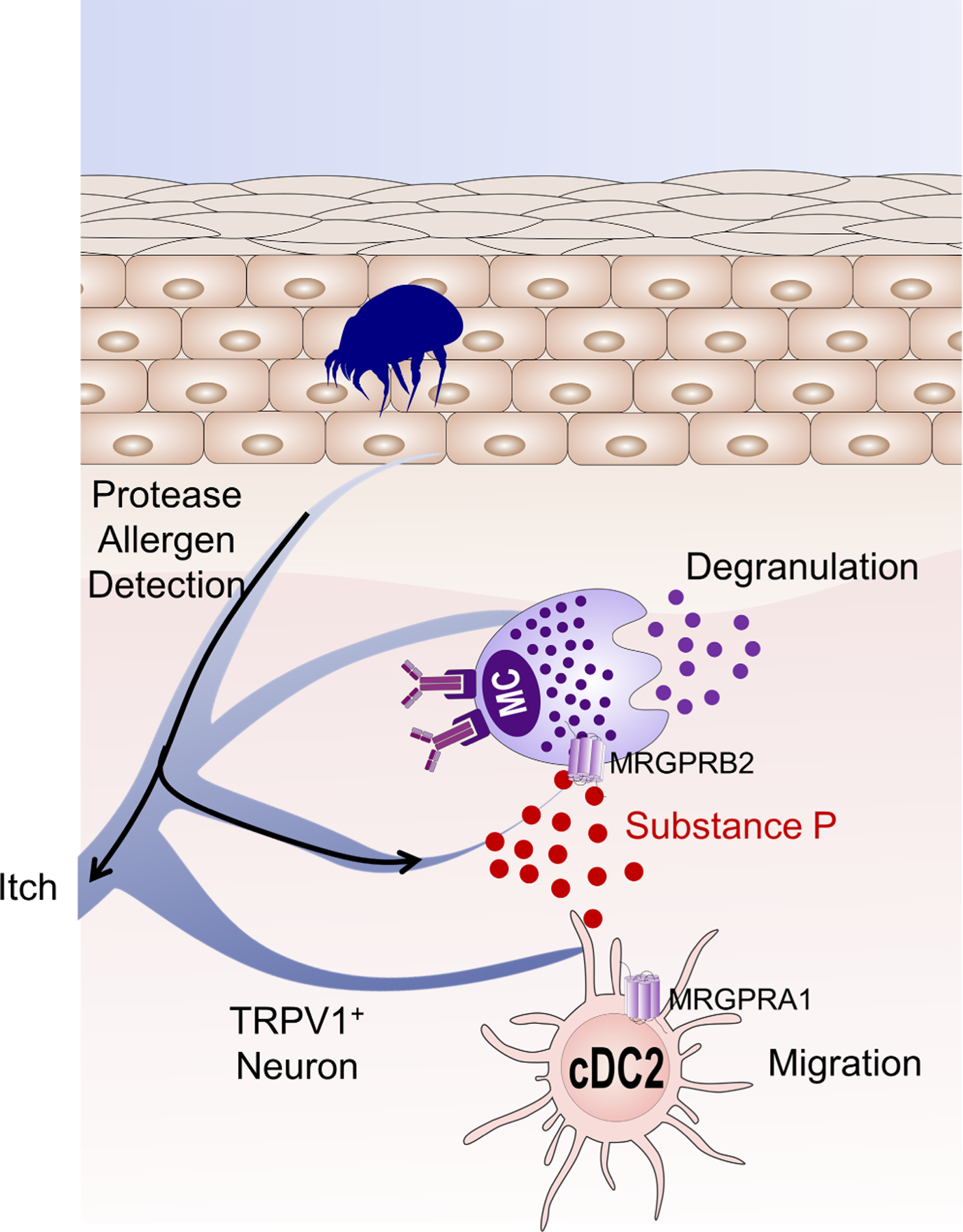
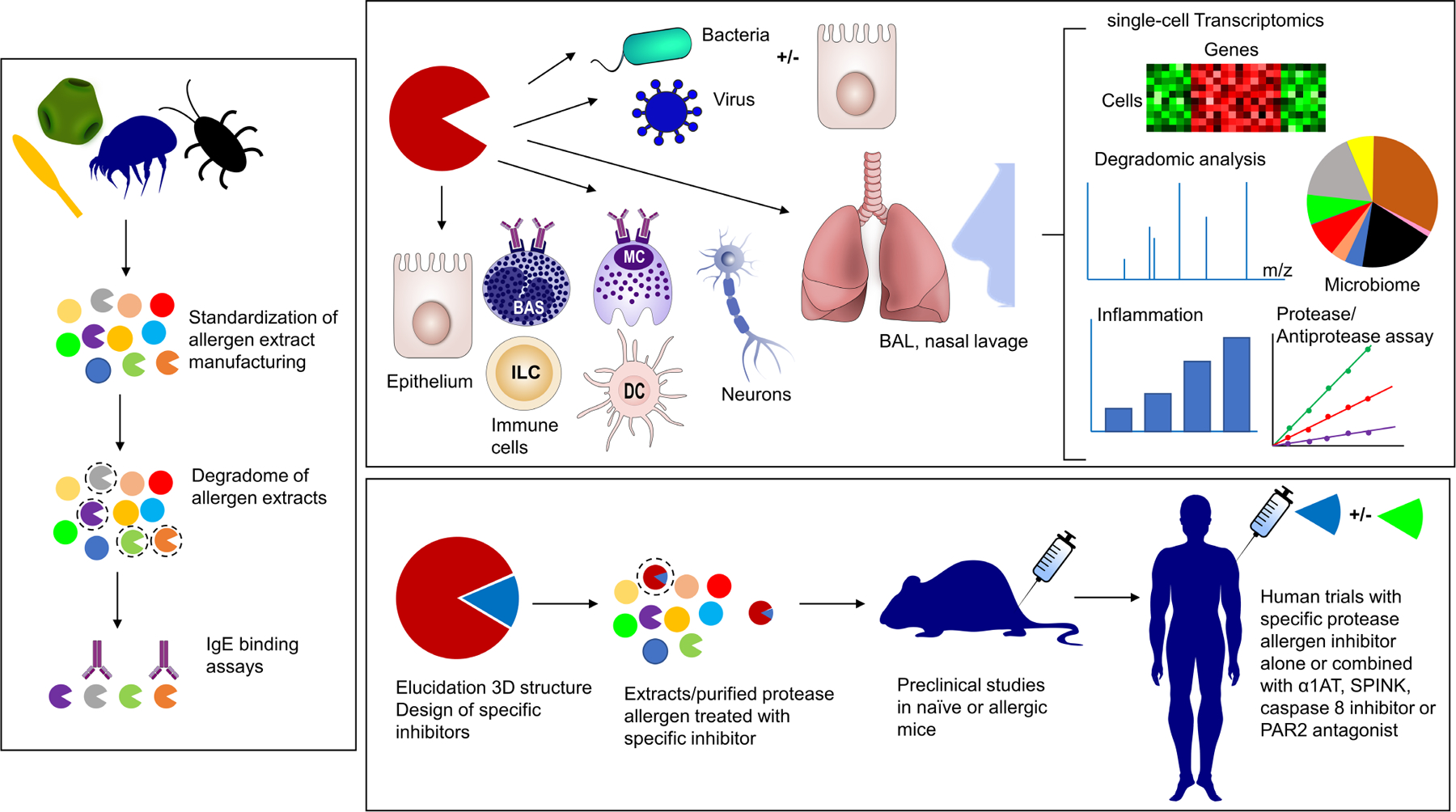
Tremendous progress in the last few years has been made to explain how seemingly harmless environmental proteins from different origins can induce potent Th2-biased inflammatory responses. Convergent findings have shown the key roles of allergens displaying proteolytic activity in the initiation and progression of the allergic response. Through their propensity to activate IgE-independent inflammatory pathways, certain allergenic proteases are now considered as initiators for sensitization to themselves and to non-protease allergens. The protease allergens degrade junctional proteins of keratinocytes or airway epithelium to facilitate allergen delivery across the epithelial barrier and their subsequent uptake by antigen-presenting cells. Epithelial injuries mediated by these proteases together with their sensing by protease-activated receptors (PARs) elicit potent inflammatory responses resulting in the release of pro-Th2 cytokines (IL-6, IL-25, IL-1β, TSLP) and danger-associated molecular patterns (DAMPs; IL-33, ATP, uric acid). Recently, protease allergens were shown to cleave the protease sensor domain of IL-33 to produce a super-active form of the alarmin. At the same time, proteolytic cleavage of fibrinogen can trigger TLR4 signaling, and cleavage of various cell surface receptors further shape the Th2 polarization. Remarkably, the sensing of protease allergens by nociceptive neurons can represent a primary step in the development of the allergic response. The goal of this review is to highlight the multiple innate immune mechanisms triggered by protease allergens that converge to initiate the allergic response.
Keywords: IL-33; protease allergen; epithelial cell; protease-activated receptor; sensory neuron.
© 2023 European Academy of Allergy and Clinical Immunology and John Wiley & Sons Ltd.
Conflict of interest statement
C.L.S is a paid consultant (expert witness) for Bayer and Merck. C.R. and J.Z. are inventors on patents granted in multiple territories concerning allergens, protease inhibitor design and composition of matter, and medical uses thereof. St George’s, University of London is beneficial owner of this patent estate. C.R. is co-founder and holds equity in Aldezon Ltd. M.E.R. is a consultant for Pulm One, Spoon Guru, ClostraBio, Serpin Pharm, Allakos, Celldex, Nextstone One, Bristol Myers Squibb, Astra Zeneca, Ellodi Pharma, GlaxoSmith Kline, Regeneron/Sanofi, Revolo Biotherapeutics, and Guidepoint and has an equity interest in the first seven listed, and royalties from reslizumab (Teva Pharmaceuticals), PEESSv2 (Mapi Research Trust) and UpToDate. M.E.R. is an inventor of patents owned by Cincinnati Children’s Hospital. The remaining authors declare that they have no conflicts of interest.
Figures





Similar articles
-
The HDM allergen orchestra and its cysteine protease maestro: Stimulators of kaleidoscopic innate immune responses.Mol Immunol. 2023 Apr;156:48-60. doi: 10.1016/j.molimm.2023.03.002. Epub 2023 Mar 6. Mol Immunol. 2023. PMID: 36889186 Review.
-
Fibrinogen cleavage products and Toll-like receptor 4 promote the generation of programmed cell death 1 ligand 2-positive dendritic cells in allergic asthma.J Allergy Clin Immunol. 2018 Aug;142(2):530-541.e6. doi: 10.1016/j.jaci.2017.09.019. Epub 2017 Oct 14. J Allergy Clin Immunol. 2018. PMID: 29038008
-
Antagonism of Protease Activated Receptor-2 by GB88 Reduces Inflammation Triggered by Protease Allergen Tyr-p3.Front Immunol. 2021 Sep 8;12:557433. doi: 10.3389/fimmu.2021.557433. eCollection 2021. Front Immunol. 2021. PMID: 34566947 Free PMC article.
-
Allergens produce serine proteases-dependent distinct release of metabolite DAMPs in human bronchial epithelial cells.Clin Exp Allergy. 2018 Feb;48(2):156-166. doi: 10.1111/cea.13071. Epub 2017 Dec 28. Clin Exp Allergy. 2018. PMID: 29210131
-
Sensing of protease activity as a triggering mechanism of Th2 cell immunity and allergic disease.Front Allergy. 2023 Sep 21;4:1265049. doi: 10.3389/falgy.2023.1265049. eCollection 2023. Front Allergy. 2023. PMID: 37810200 Free PMC article. Review.
Cited by
-
Editorial: The regulation of allergic responses by proteolysis: from protease allergens to host proteases modulation.Front Allergy. 2024 Aug 12;5:1469718. doi: 10.3389/falgy.2024.1469718. eCollection 2024. Front Allergy. 2024. PMID: 39188990 Free PMC article. No abstract available.
-
The New Paradigm: The Role of Proteins and Triggers in the Evolution of Allergic Asthma.Int J Mol Sci. 2024 May 25;25(11):5747. doi: 10.3390/ijms25115747. Int J Mol Sci. 2024. PMID: 38891935 Free PMC article. Review.
-
Epithelial-immune interactions govern type 2 immunity at barrier surfaces.Curr Opin Immunol. 2024 Dec;91:102501. doi: 10.1016/j.coi.2024.102501. Epub 2024 Nov 10. Curr Opin Immunol. 2024. PMID: 39522453 Review.
-
Activation of the Coagulation Cascade as a Universal Danger Sign.Curr Issues Mol Biol. 2025 Feb 9;47(2):108. doi: 10.3390/cimb47020108. Curr Issues Mol Biol. 2025. PMID: 39996829 Free PMC article. Review.
-
Polymerised mite allergoids with glutaraldehyde reduce proteolytic activity and enhance the stability of allergen mixtures: a proof of concept with grass mixtures.Front Allergy. 2025 Feb 25;6:1557650. doi: 10.3389/falgy.2025.1557650. eCollection 2025. Front Allergy. 2025. PMID: 40071131 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
