

High Spectral and Temporal Acuity in Primary Auditory Cortex of Awake Cats
- PMID: 36795196
- PMCID: PMC10121981
- DOI: 10.1007/s10162-023-00890-6
High Spectral and Temporal Acuity in Primary Auditory Cortex of Awake Cats
Abstract
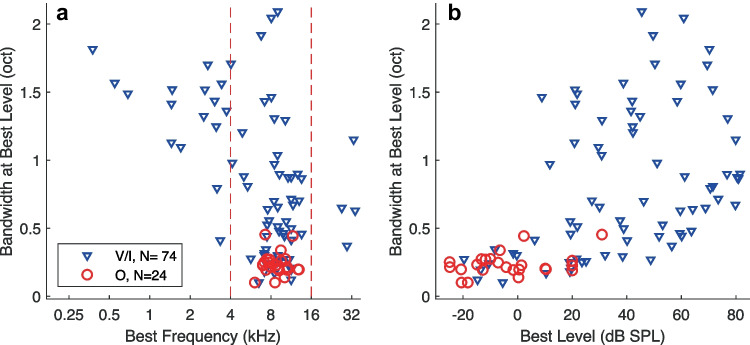
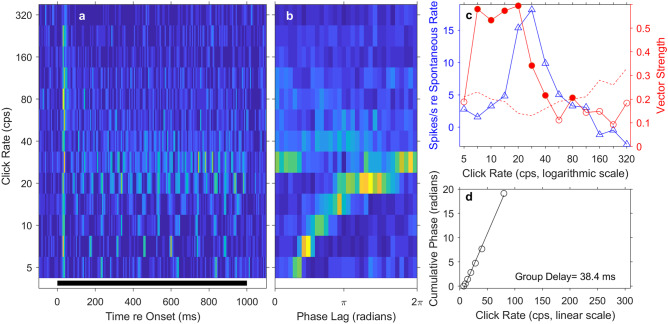
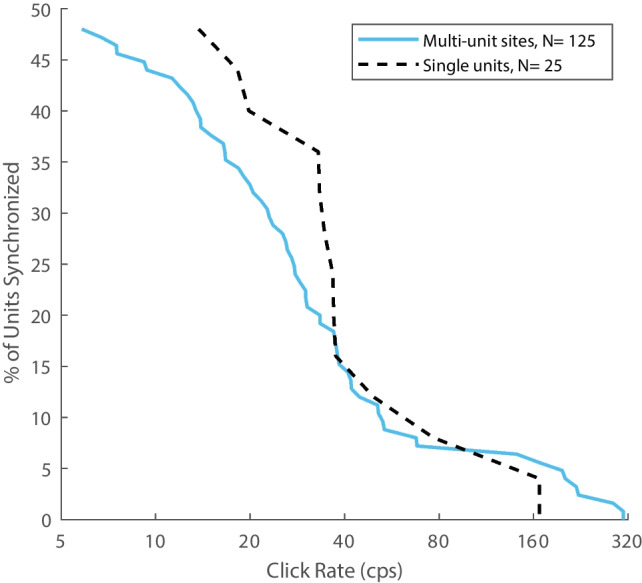
Most accounts of single- and multi-unit responses in auditory cortex under anesthetized conditions have emphasized V-shaped frequency tuning curves and low-pass sensitivity to rates of repeated sounds. In contrast, single-unit recordings in awake marmosets also show I-shaped and O-shaped response areas having restricted tuning to frequency and (for O units) sound level. That preparation also demonstrates synchrony to moderate click rates and representation of higher click rates by spike rates of non-synchronized tonic responses, neither of which are commonly seen in anesthetized conditions. The spectral and temporal representation observed in the marmoset might reflect special adaptations of that species, might be due to single- rather than multi-unit recording, or might indicate characteristics of awake-versus-anesthetized recording conditions. We studied spectral and temporal representation in the primary auditory cortex of alert cats. We observed V-, I-, and O-shaped response areas like those demonstrated in awake marmosets. Neurons could synchronize to click trains at rates about an octave higher than is usually seen with anesthesia. Representations of click rates by rates of non-synchronized tonic responses exhibited dynamic ranges that covered the entire range of tested click rates. The observation of these spectral and temporal representations in cats demonstrates that they are not unique to primates and, indeed, might be widespread among mammalian species. Moreover, we observed no significant difference in stimulus representation between single- and multi-unit recordings. It appears that the principal factor that has hindered observations of high spectral and temporal acuity in the auditory cortex has been the use of general anesthesia.
Keywords: Auditory cortex; Auditory filters; Frequency response area; Non-monotonic; Non-synchronized; Phase locking; Temporal representation; Tuning curve.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures










References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
