

This is a preprint.
Multiplexed Subspaces Route Neural Activity Across Brain-wide Networks
- PMID: 36798411
- PMCID: PMC9934668
- DOI: 10.1101/2023.02.08.527772
Multiplexed Subspaces Route Neural Activity Across Brain-wide Networks
Update in
-
Multiplexed subspaces route neural activity across brain-wide networks.Nat Commun. 2025 Apr 9;16(1):3359. doi: 10.1038/s41467-025-58698-2. Nat Commun. 2025. PMID: 40204762 Free PMC article.
Abstract
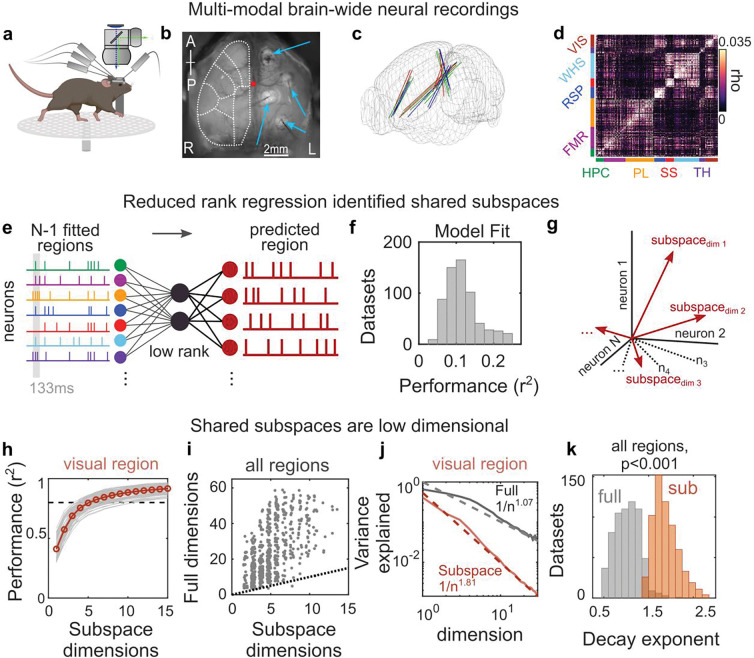
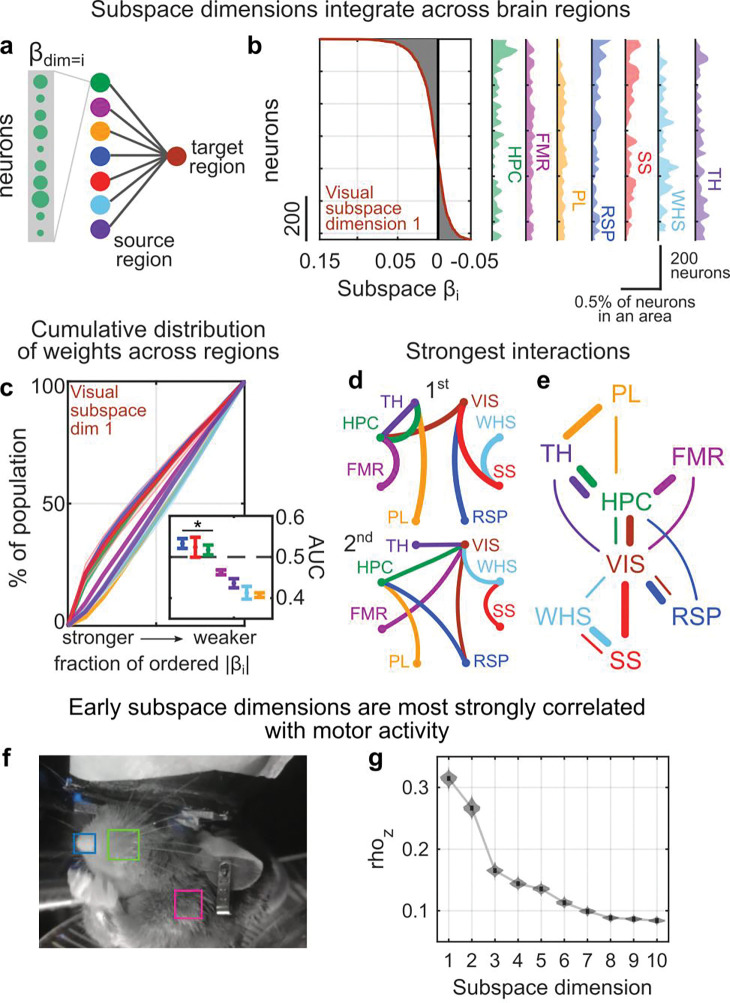
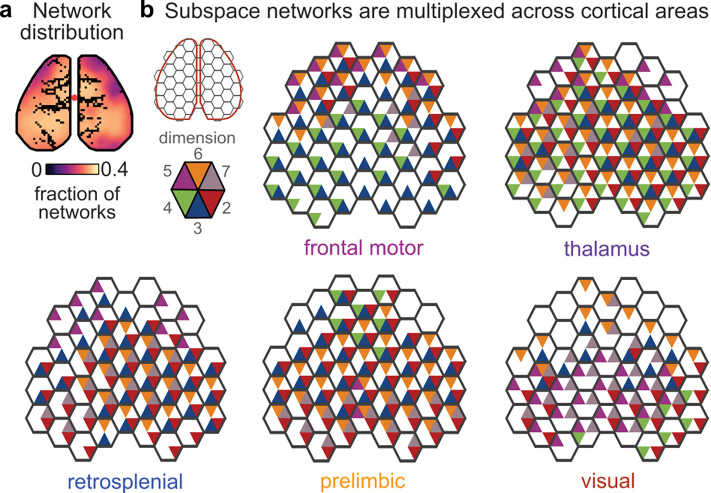
Cognition is flexible. Behaviors can change on a moment-by-moment basis. Such flexibility is thought to rely on the brain's ability to route information through different networks of brain regions in order to support different cognitive computations. However, the mechanisms that determine which network of brain regions is engaged are unknown. To address this, we combined cortex-wide calcium imaging with high-density electrophysiological recordings in eight cortical and subcortical regions of mice. Different dimensions within the population activity of each brain region were functionally connected with different cortex-wide 'subspace networks' of regions. These subspace networks were multiplexed, allowing a brain region to simultaneously interact with multiple independent, yet overlapping, networks. Alignment of neural activity within a region to a specific subspace network dimension predicted how neural activity propagated between regions. Thus, changing the geometry of the neural representation within a brain region could be a mechanism to selectively engage different brain-wide networks to support cognitive flexibility.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures



References
-
- MacDowell C. J., Briones B. A., Lenzi M. J., Gustison M. L. & Buschman T. J. Variability in sampling of cortex-wide neural dynamics explains individual differences in functional connectivity and behavioral phenotype. 2022.01.24.477572 Preprint at 10.1101/2022.01.24.477572 (2022). - DOI
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources