

Tripartite interactions between filamentous Pf4 bacteriophage, Pseudomonas aeruginosa, and bacterivorous nematodes
- PMID: 36800381
- PMCID: PMC9980816
- DOI: 10.1371/journal.ppat.1010925
Tripartite interactions between filamentous Pf4 bacteriophage, Pseudomonas aeruginosa, and bacterivorous nematodes
Abstract
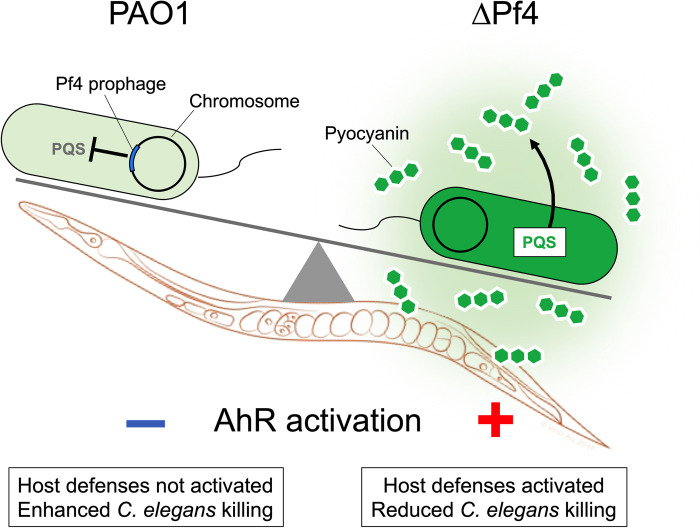
The opportunistic pathogen Pseudomonas aeruginosa PAO1 is infected by the filamentous bacteriophage Pf4. Pf4 virions promote biofilm formation, protect bacteria from antibiotics, and modulate animal immune responses in ways that promote infection. Furthermore, strains cured of their Pf4 infection (ΔPf4) are less virulent in animal models of infection. Consistently, we find that strain ΔPf4 is less virulent in a Caenorhabditis elegans nematode infection model. However, our data indicate that PQS quorum sensing is activated and production of the pigment pyocyanin, a potent virulence factor, is enhanced in strain ΔPf4. The reduced virulence of ΔPf4 despite high levels of pyocyanin production may be explained by our finding that C. elegans mutants unable to sense bacterial pigments through the aryl hydrocarbon receptor are more susceptible to ΔPf4 infection compared to wild-type C. elegans. Collectively, our data support a model where suppression of quorum-regulated virulence factors by Pf4 allows P. aeruginosa to evade detection by innate host immune responses.
Copyright: © 2023 Schwartzkopf et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures






References
-
- Secor PR, Burgener EB, Kinnersley M, Jennings LK, Roman-Cruz V, Popescu M, et al.. Pf Bacteriophage and Their Impact on Pseudomonas Virulence, Mammalian Immunity, and Chronic Infections. Front Immunol. 2020;11:244. Epub 2020/03/11. doi: 10.3389/fimmu.2020.00244 ; PubMed Central PMCID: PMC7047154. - DOI - PMC - PubMed
-
- Rakonjac J, Bennett NJ, Spagnuolo J, Gagic D, Russel M. Filamentous Bacteriophage: Biology, Phage Display and Nanotechnology Applications. Curr Issues Mol Biol. 2011;13(2):51–76. Epub 2011/04/20. . - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
