

Intraluminal pressure elevates intracellular calcium and contracts CNS pericytes: Role of voltage-dependent calcium channels
- PMID: 36802432
- PMCID: PMC9992766
- DOI: 10.1073/pnas.2216421120
Intraluminal pressure elevates intracellular calcium and contracts CNS pericytes: Role of voltage-dependent calcium channels
Abstract
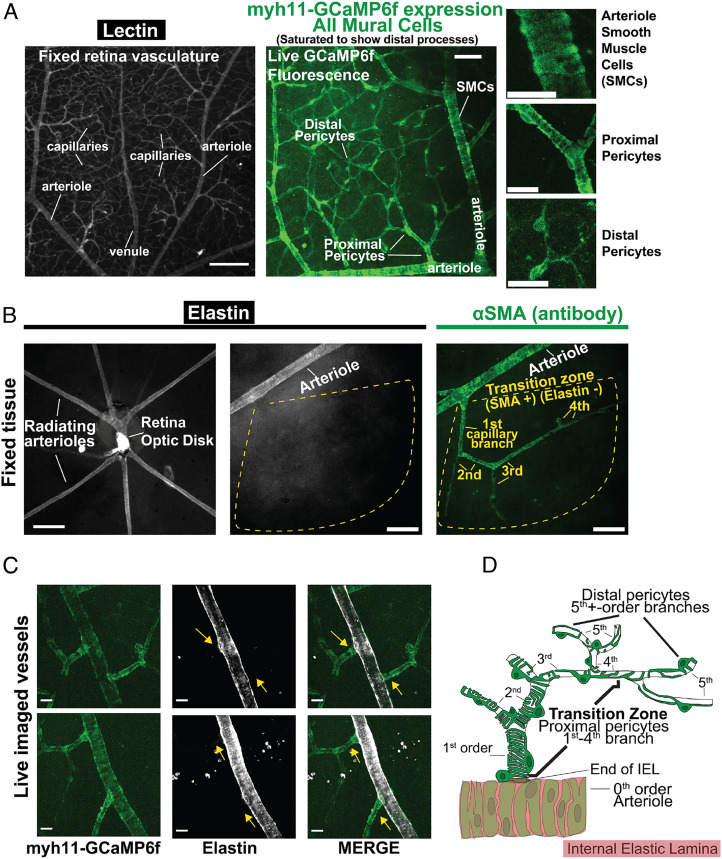
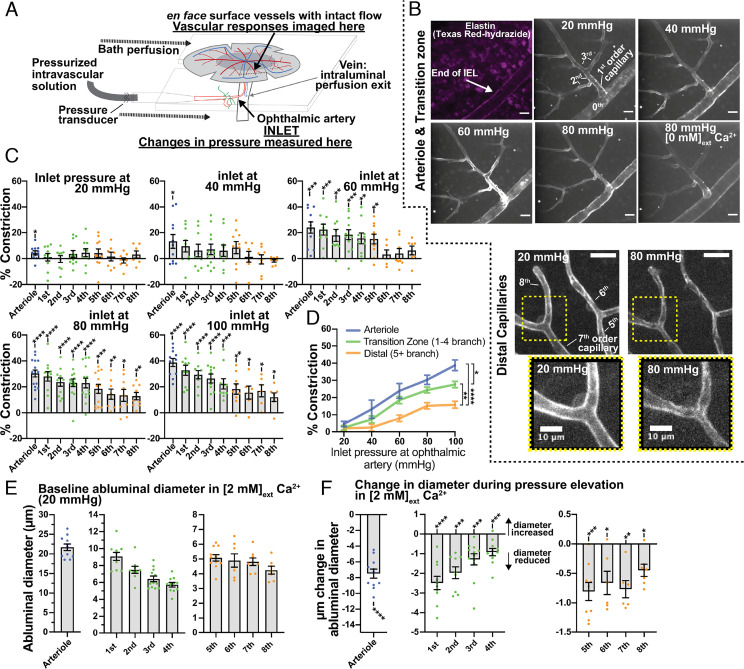
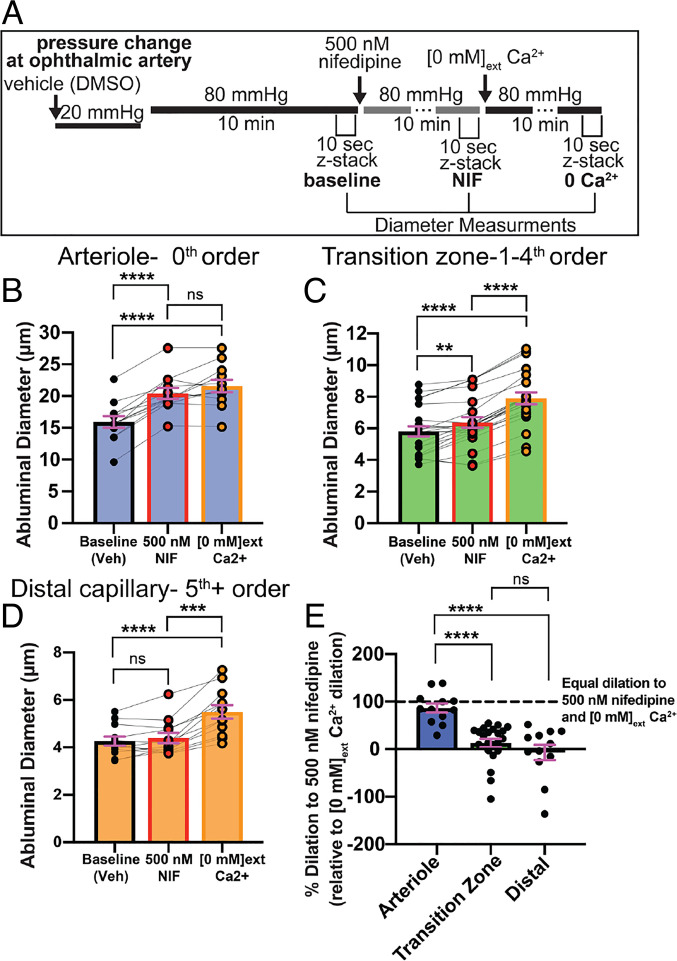
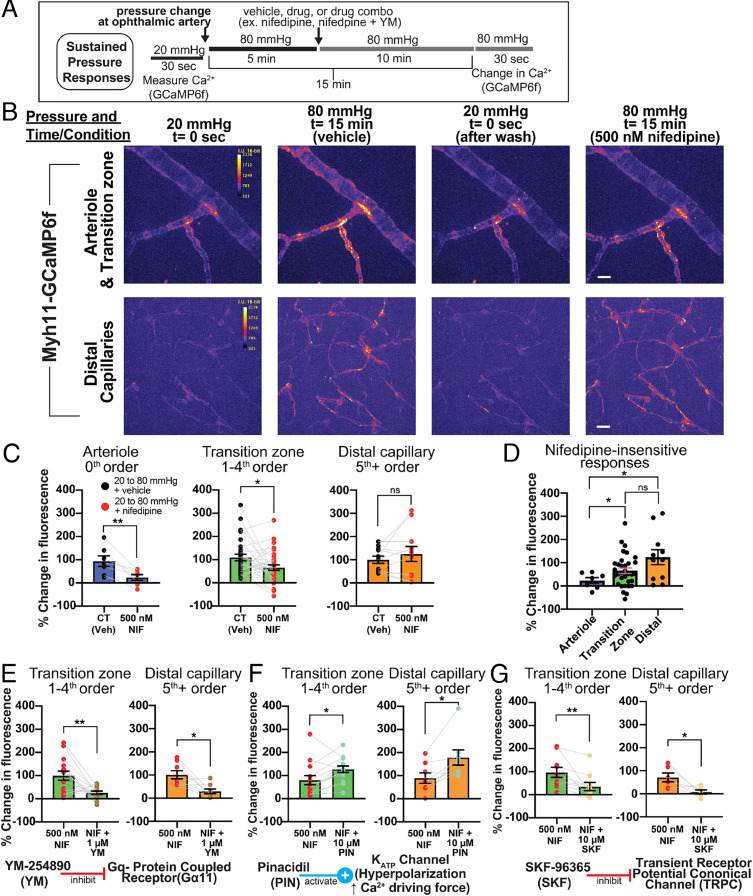
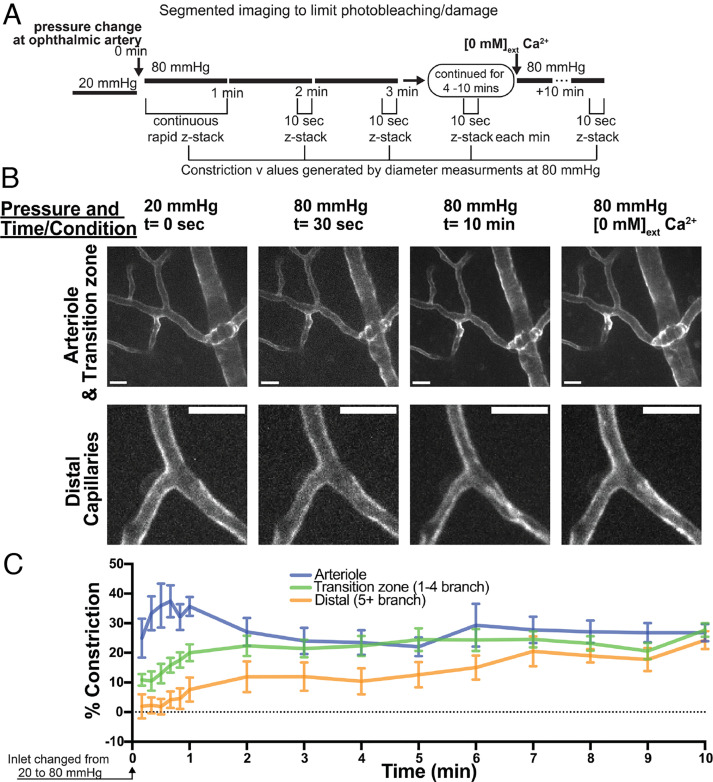
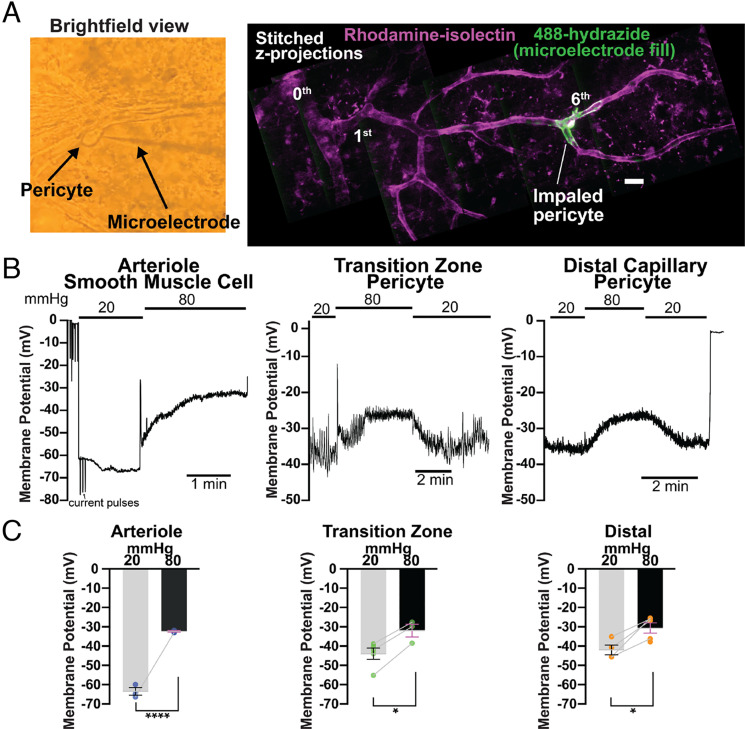
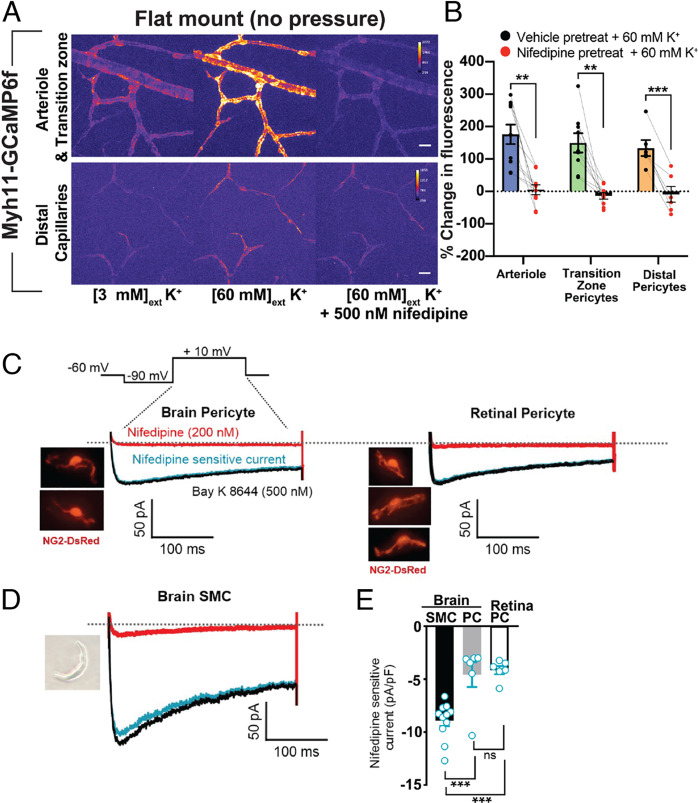
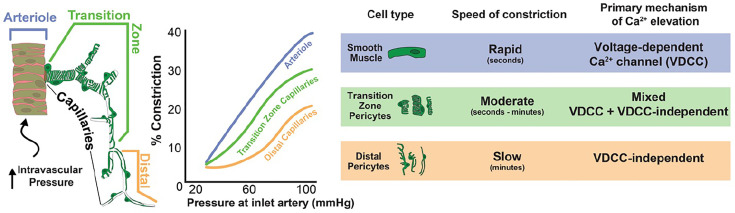
Arteriolar smooth muscle cells (SMCs) and capillary pericytes dynamically regulate blood flow in the central nervous system in the face of fluctuating perfusion pressures. Pressure-induced depolarization and Ca2+ elevation provide a mechanism for regulation of SMC contraction, but whether pericytes participate in pressure-induced changes in blood flow remains unknown. Here, utilizing a pressurized whole-retina preparation, we found that increases in intraluminal pressure in the physiological range induce contraction of both dynamically contractile pericytes in the arteriole-proximate transition zone and distal pericytes of the capillary bed. We found that the contractile response to pressure elevation was slower in distal pericytes than in transition zone pericytes and arteriolar SMCs. Pressure-evoked elevation of cytosolic Ca2+ and contractile responses in SMCs were dependent on voltage-dependent Ca2+ channel (VDCC) activity. In contrast, Ca2+ elevation and contractile responses were partially dependent on VDCC activity in transition zone pericytes and independent of VDCC activity in distal pericytes. In both transition zone and distal pericytes, membrane potential at low inlet pressure (20 mmHg) was approximately -40 mV and was depolarized to approximately -30 mV by an increase in pressure to 80 mmHg. The magnitude of whole-cell VDCC currents in freshly isolated pericytes was approximately half that measured in isolated SMCs. Collectively, these results indicate a loss of VDCC involvement in pressure-induced constriction along the arteriole-capillary continuum. They further suggest that alternative mechanisms and kinetics of Ca2+ elevation, contractility, and blood flow regulation exist in central nervous system capillary networks, distinguishing them from neighboring arterioles.
Keywords: autoregulation; cerebral blood flow; pericytes; vascular tone.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
