

Directed differentiation of human iPSCs to functional ovarian granulosa-like cells via transcription factor overexpression
- PMID: 36803359
- PMCID: PMC9943069
- DOI: 10.7554/eLife.83291
Directed differentiation of human iPSCs to functional ovarian granulosa-like cells via transcription factor overexpression
Erratum in
-
Correction: Directed differentiation of human iPSCs to functional ovarian granulosa-like cells via transcription factor overexpression.Elife. 2023 Mar 22;12:e87987. doi: 10.7554/eLife.87987. Elife. 2023. PMID: 36947114 Free PMC article.
Abstract
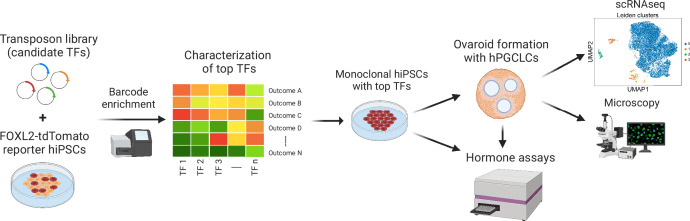
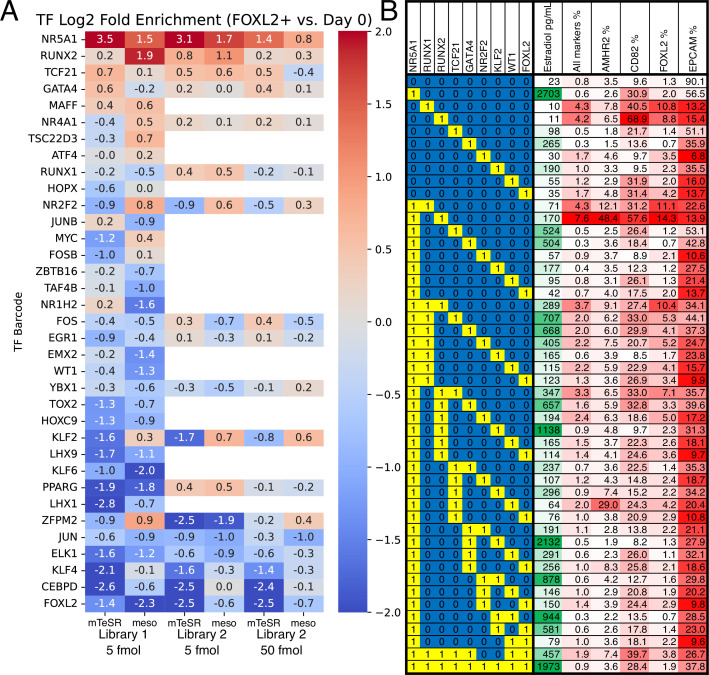
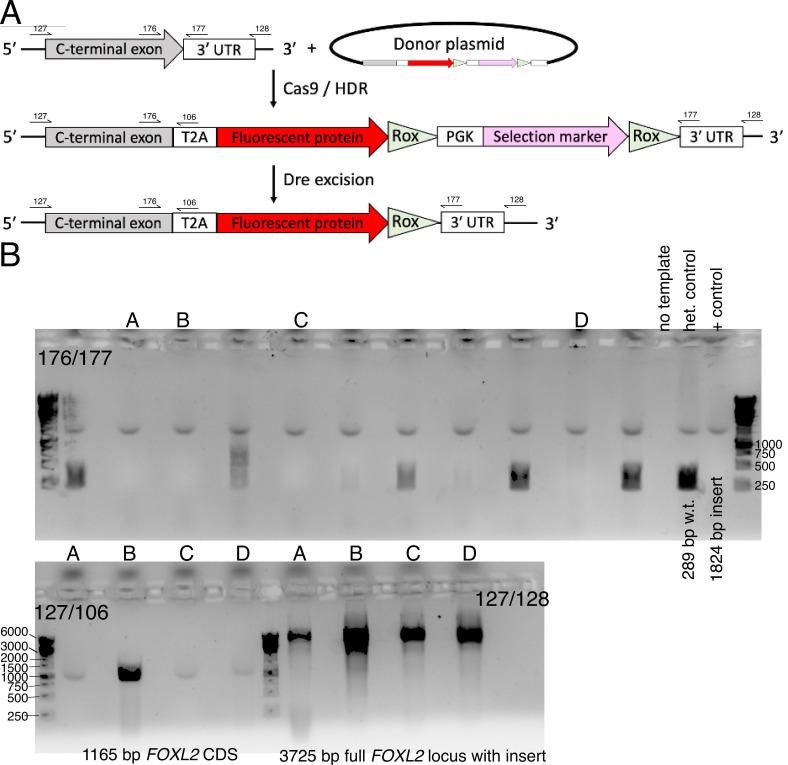
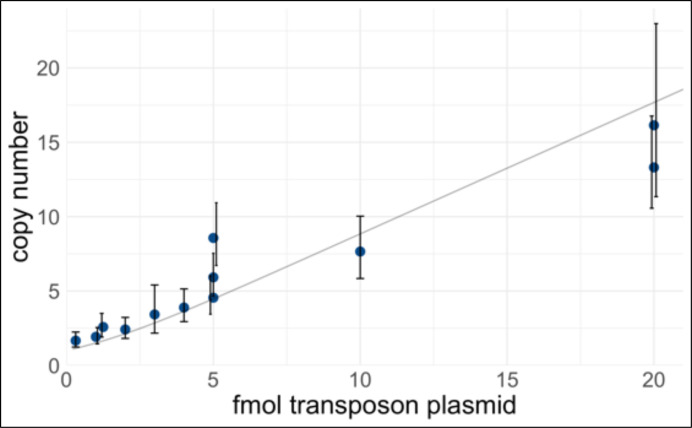
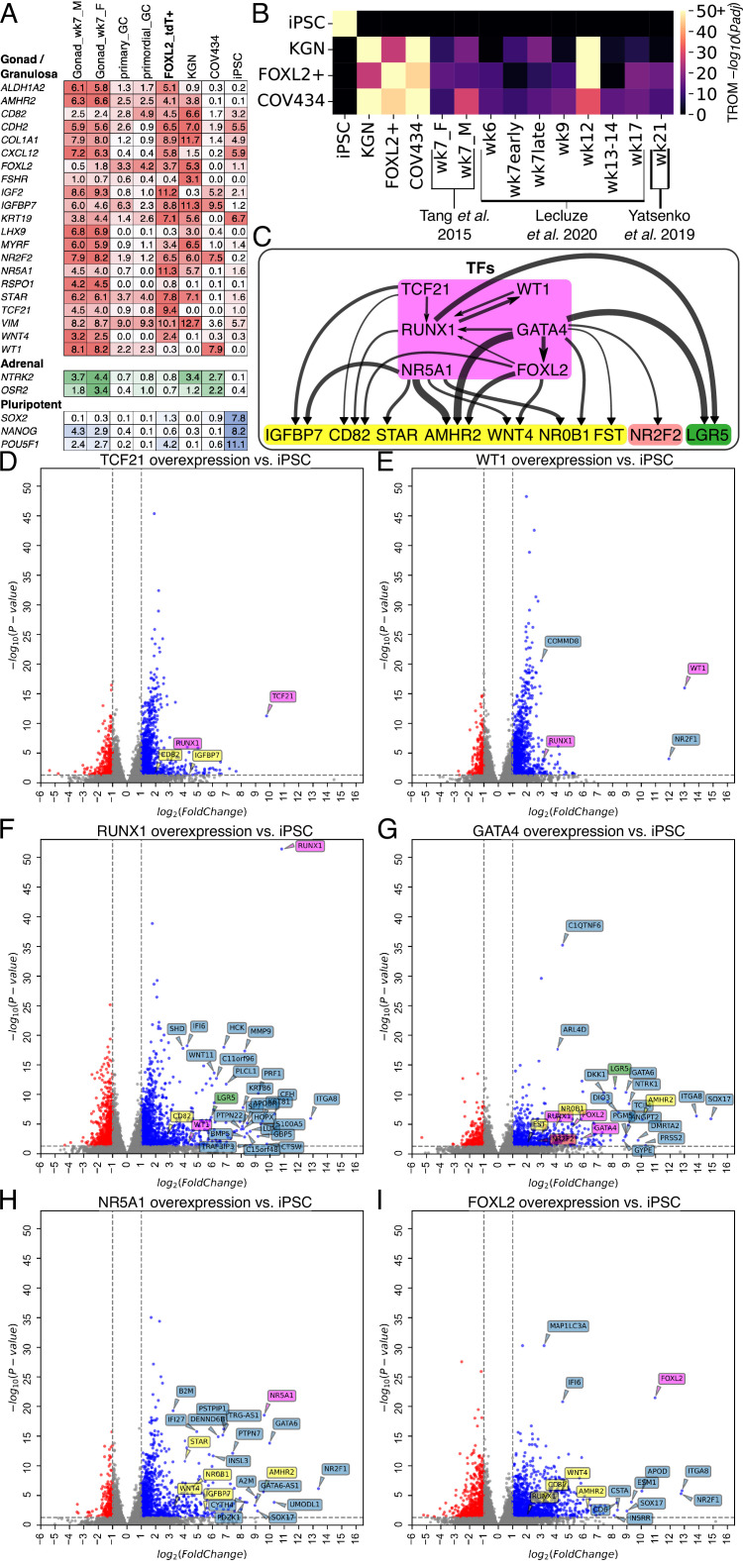
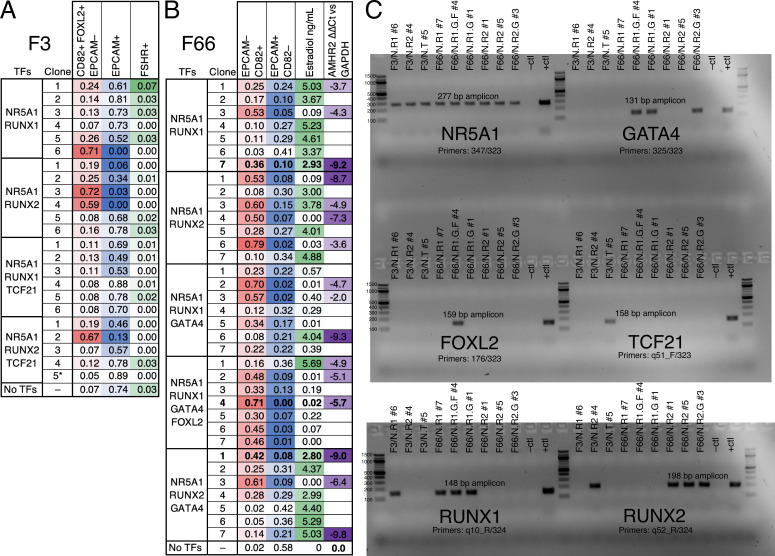
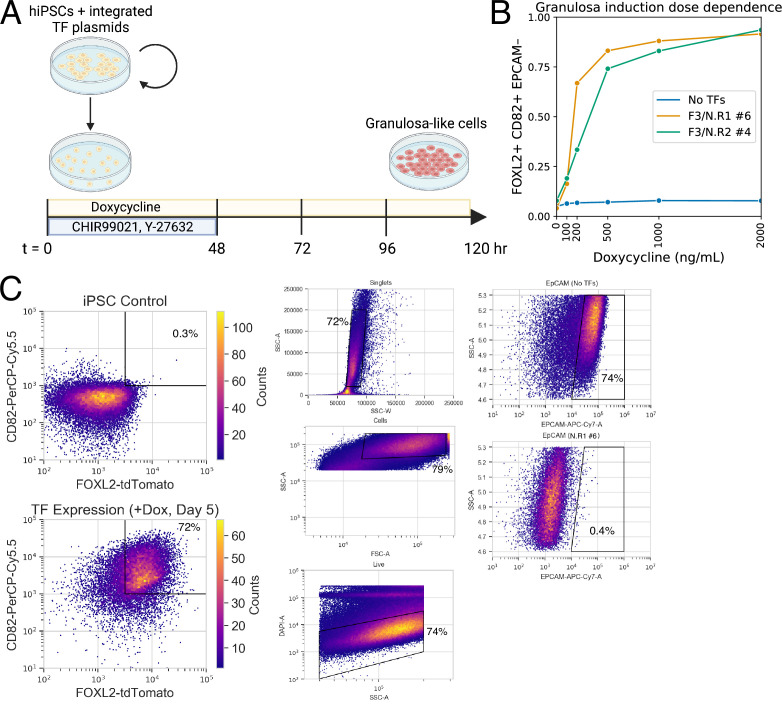
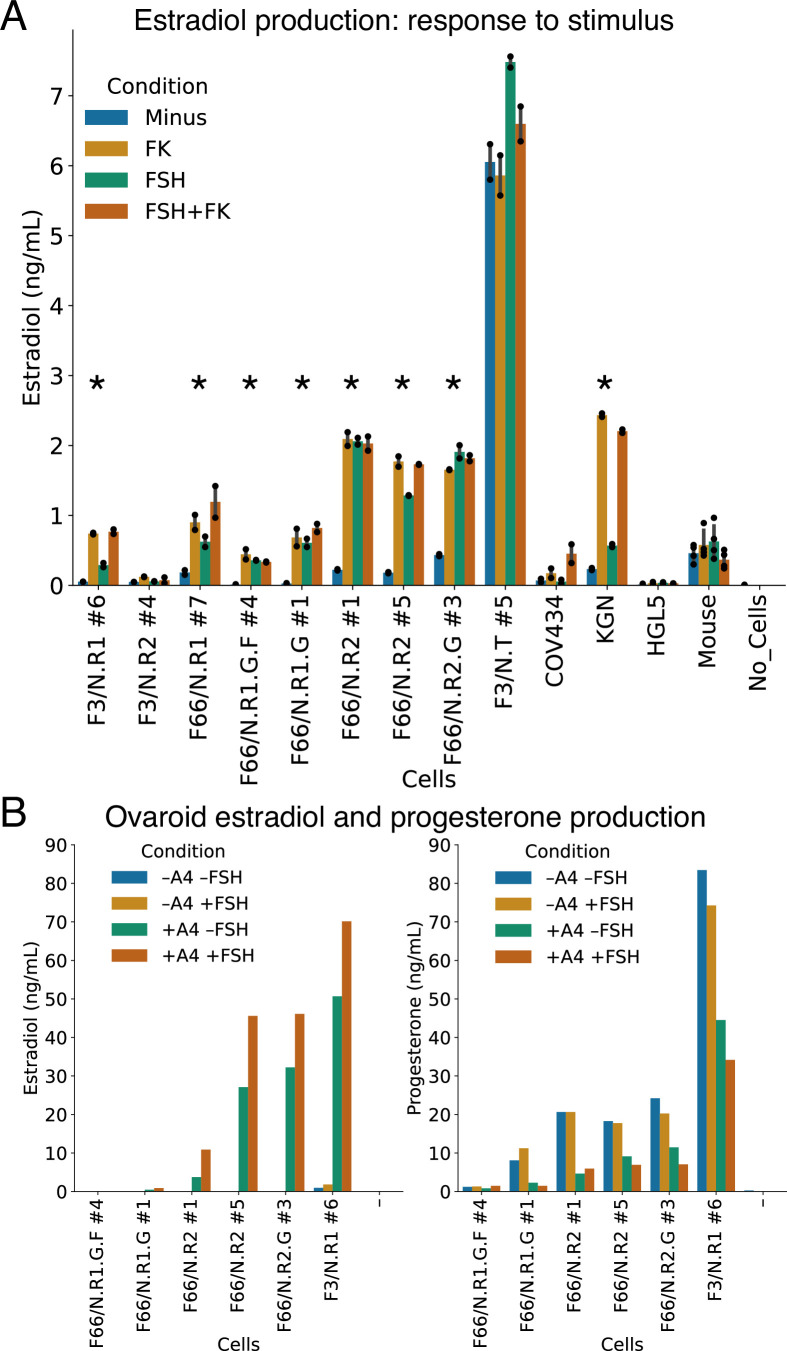
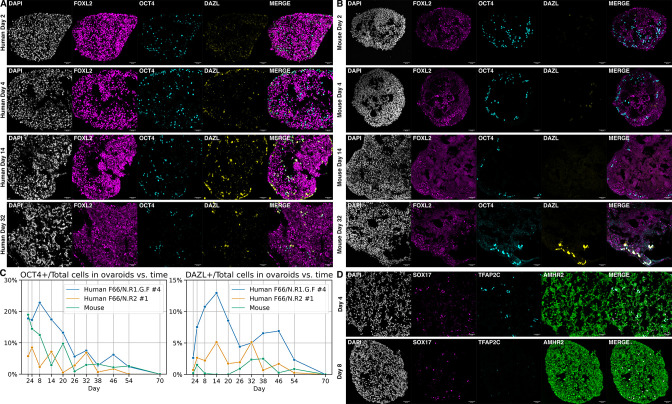
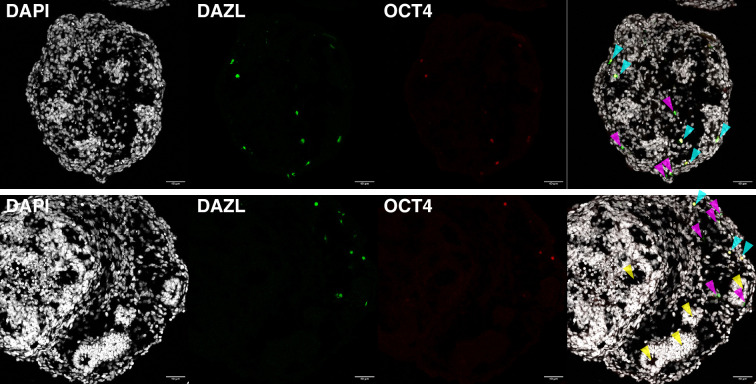
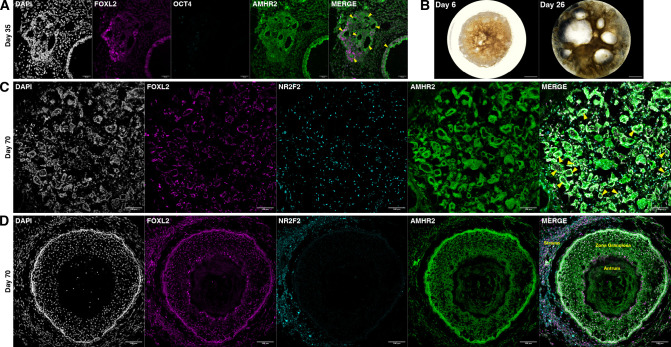
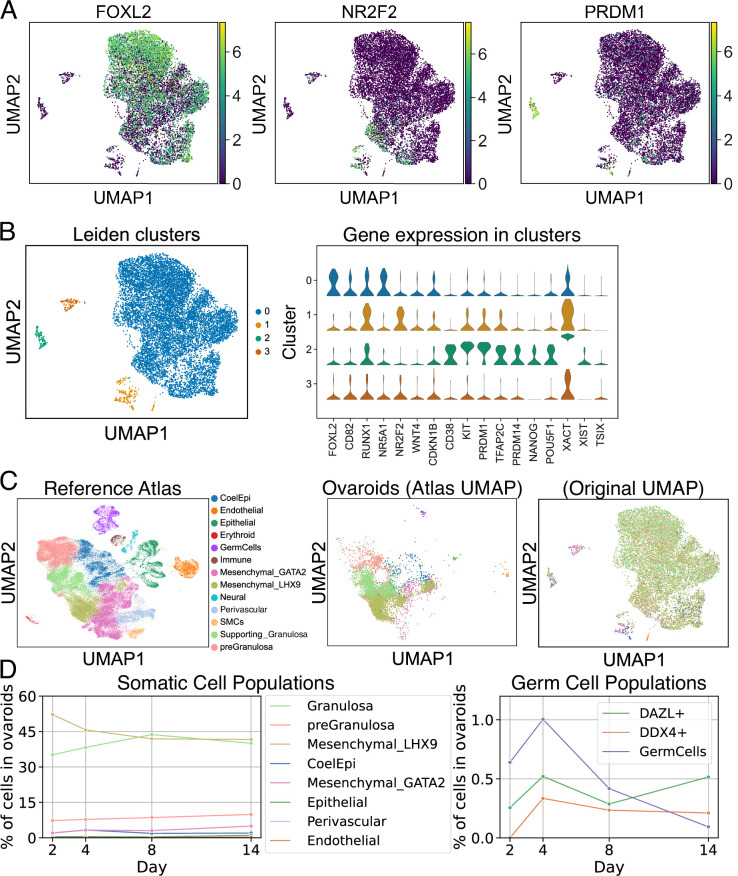
An in vitro model of human ovarian follicles would greatly benefit the study of female reproduction. Ovarian development requires the combination of germ cells and several types of somatic cells. Among these, granulosa cells play a key role in follicle formation and support for oogenesis. Whereas efficient protocols exist for generating human primordial germ cell-like cells (hPGCLCs) from human induced pluripotent stem cells (hiPSCs), a method of generating granulosa cells has been elusive. Here, we report that simultaneous overexpression of two transcription factors (TFs) can direct the differentiation of hiPSCs to granulosa-like cells. We elucidate the regulatory effects of several granulosa-related TFs and establish that overexpression of NR5A1 and either RUNX1 or RUNX2 is sufficient to generate granulosa-like cells. Our granulosa-like cells have transcriptomes similar to human fetal ovarian cells and recapitulate key ovarian phenotypes including follicle formation and steroidogenesis. When aggregated with hPGCLCs, our cells form ovary-like organoids (ovaroids) and support hPGCLC development from the premigratory to the gonadal stage as measured by induction of DAZL expression. This model system will provide unique opportunities for studying human ovarian biology and may enable the development of therapies for female reproductive health.
Keywords: developmental biology; granulosa; human; mouse; oogenesis; ovary; regenerative medicine; stem cells; transcription factors.
Plain language summary
Ovaries are responsible for forming the eggs humans and other mammals need to reproduce. Once mature, the egg cell is released into the fallopian tube where it can be potentially fertilized by a sperm. Despite their crucial role, how eggs are made in the ovary is poorly understood. This is because ovaries are hard to access, making it difficult to conduct experiments on them. To overcome this, researchers have built artificial ovaries in the laboratory using stem cells from the embryos of mice which can develop into all cell types in the adult body. By culturing these embryonic stem cells under special conditions, researchers can convert them in to the two main cell types of the developing ovary: germ cells which go on to form eggs, and granulosa cells which help eggs grow and mature. The resulting lab-grown ovary can make eggs that produce live mice when fertilized. This approach has also been applied to human induced pluripotent stem cells (iPSCs), adult human cells which have been reprogrammed to a stem-like state. While this has produced human germ cells, generating human granulosa cells has been more challenging. Here, Pierson Smela, Kramme et al. show that activating a specific set of transcription factors (proteins that switch genes on or off) in iPSCs can make them transition to granulosa cells. First, the team tested random combinations of 35 transcription factors which, based on previous literature and genetic data, were likely to play a role in the formation of granulosa cells. This led to the identification of a small number of factors that caused the human iPSCs to develop features and carry out roles seen in mature granulosa cells; this includes producing an important reproductive hormone and supporting the maturation of germ cells. Pierson Smela, Kramme et al. found that growing these granulosa-like cells together with germ cells (also generated via iPSCs) resulted in structures similar to ovarian follicles which help eggs develop. These findings could help researchers build stable systems for studying how granulosa cells behave in human ovaries. This could lead to new insights about reproductive health.
© 2023, Pierson Smela, Kramme et al.
Conflict of interest statement
MP, MK, GB, VK, ET, RK, TS is listed as an inventor for US Provisional Application No. 63/326,640, entitled 'Methods and Compositions for Producing Granulosa-Like Cells', CK is listed as an inventor for US Provisional Application No. 63/326,640, entitled 'Methods and Compositions for Producing Granulosa-Like Cells'. CK is the VP of Cell Engineering of Gameto, Inc, PF, JA, RS, ED No competing interests declared, PC is listed an an inventor for US Provisional Application No. 63/326,640, entitled 'Methods and Compositions for Producing Granulosa-Like Cells'. PC is a co-founder and scientific advisor to Gameto, Inc, GC is listed as an inventor for US Provisional Application No. 63/326,640, entitled 'Methods and Compositions for Producing Granulosa-Like Cells'. GMC serves on the scientific advisory board of Gameto, Inc, Colossal Biosciences, and GCTx
Figures
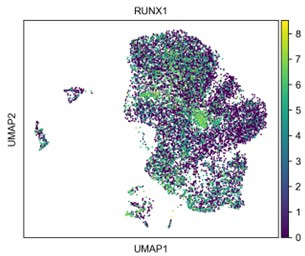
References
-
- Brixi G, Tysinger E, Smela MP, Chatterjee P. Directed differentiation of human ipscs to functional ovarian granulosa-like cells via transcription factor overexpression. swh:1:rev:3c650290779db376c4d1f3a14960b08b17ae5561Software Heritage. 2023 https://archive.softwareheritage.org/swh:1:dir:23c3e0d1110d90ccffcd2d8ef... - PubMed
-
- Chen H-H, Welling M, Bloch DB, Muñoz J, Mientjes E, Chen X, Tramp C, Wu J, Yabuuchi A, Chou Y-F, Buecker C, Krainer A, Willemsen R, Heck AJ, Geijsen N. Dazl limits pluripotency, differentiation, and apoptosis in developing primordial germ cells. Stem Cell Reports. 2014;3:892–904. doi: 10.1016/j.stemcr.2014.09.003. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
