

HIV rapidly targets a diverse pool of CD4+ T cells to establish productive and latent infections
- PMID: 36804957
- PMCID: PMC10023508
- DOI: 10.1016/j.immuni.2023.01.030
HIV rapidly targets a diverse pool of CD4+ T cells to establish productive and latent infections
Abstract
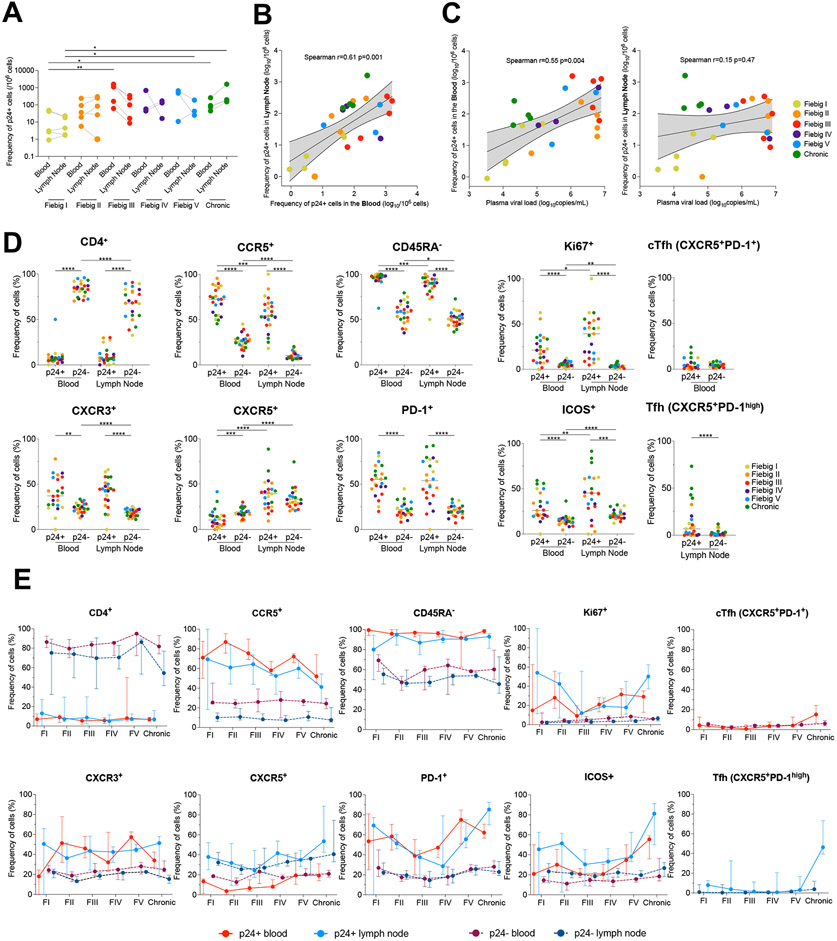
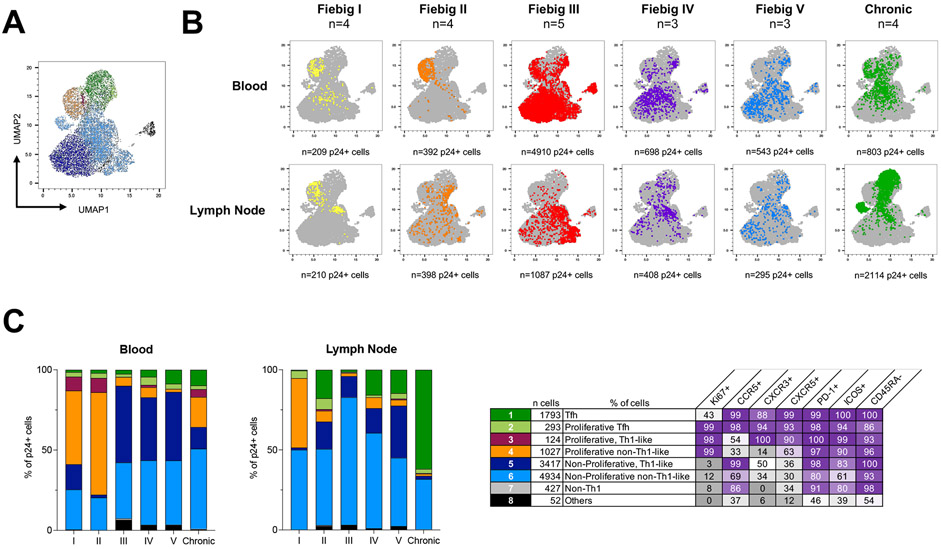
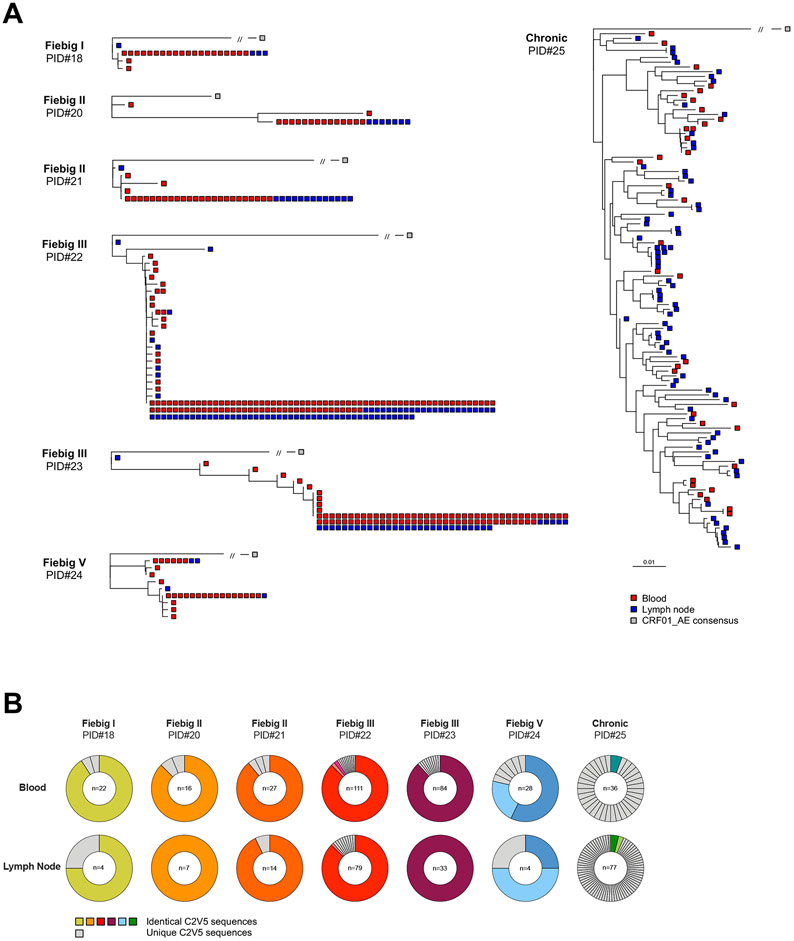
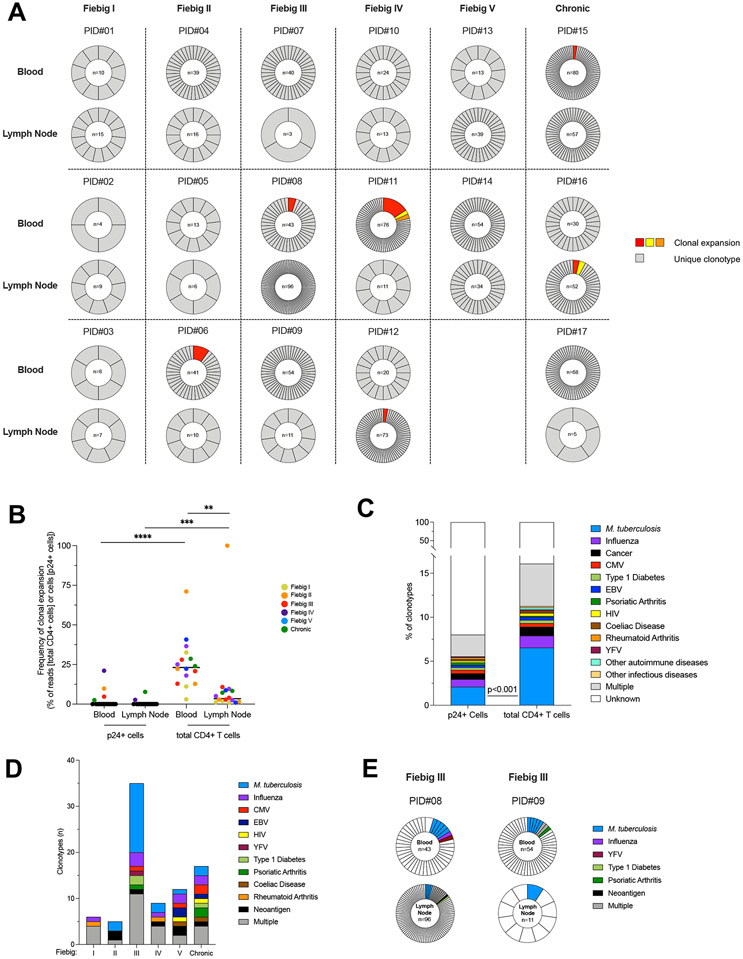
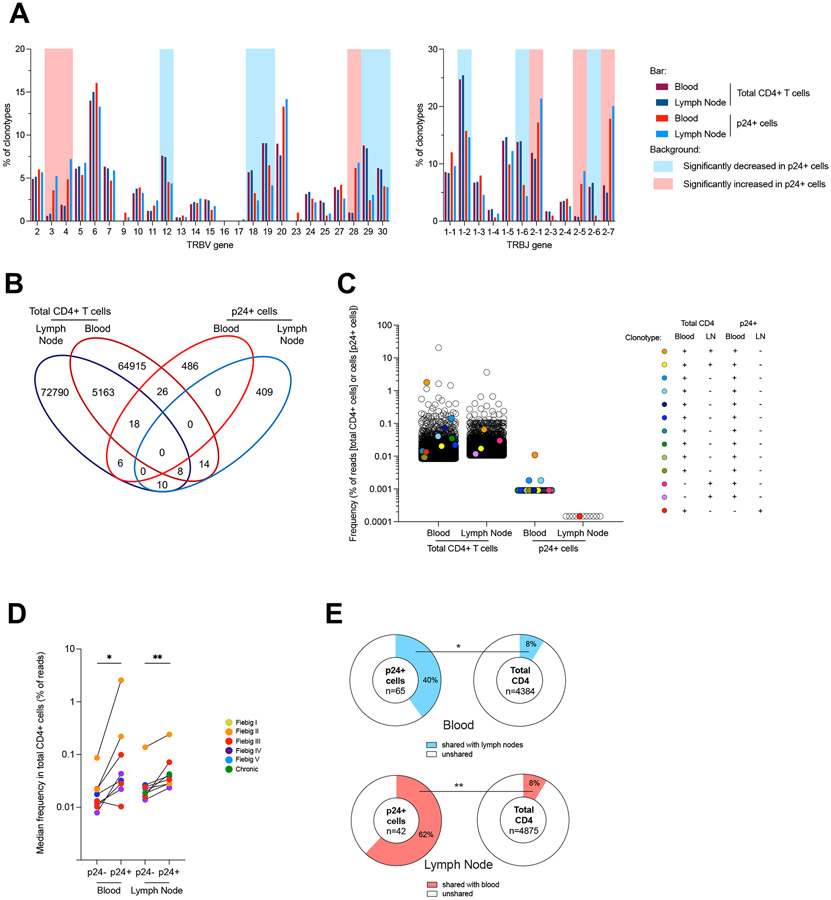
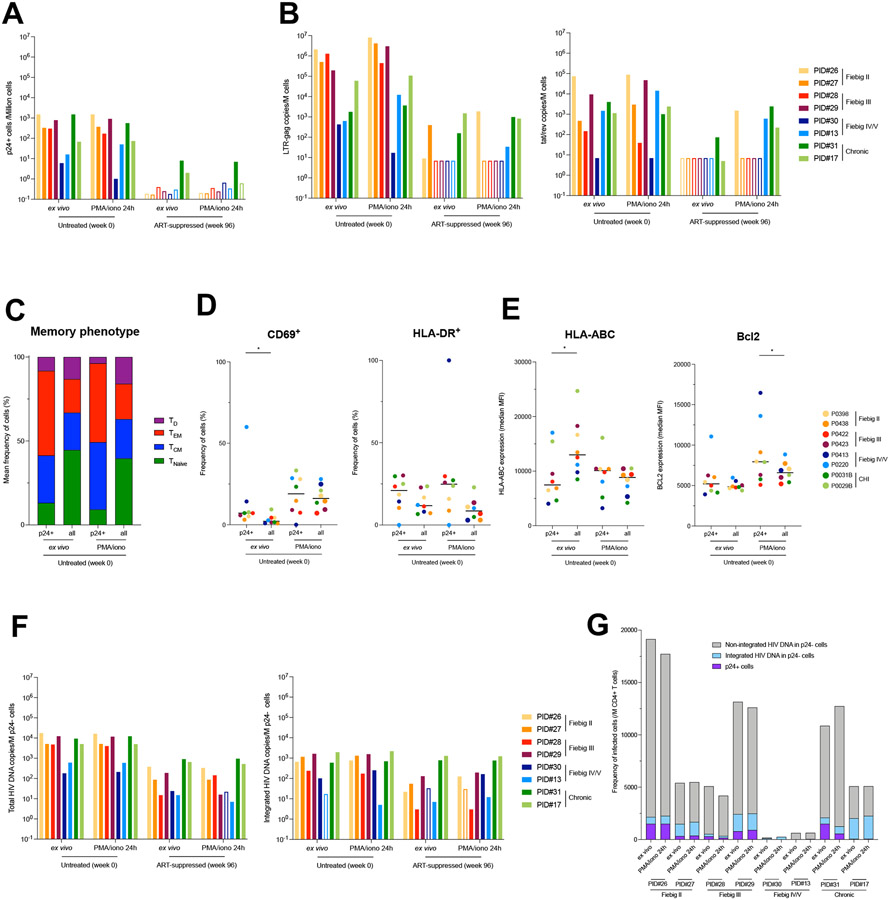
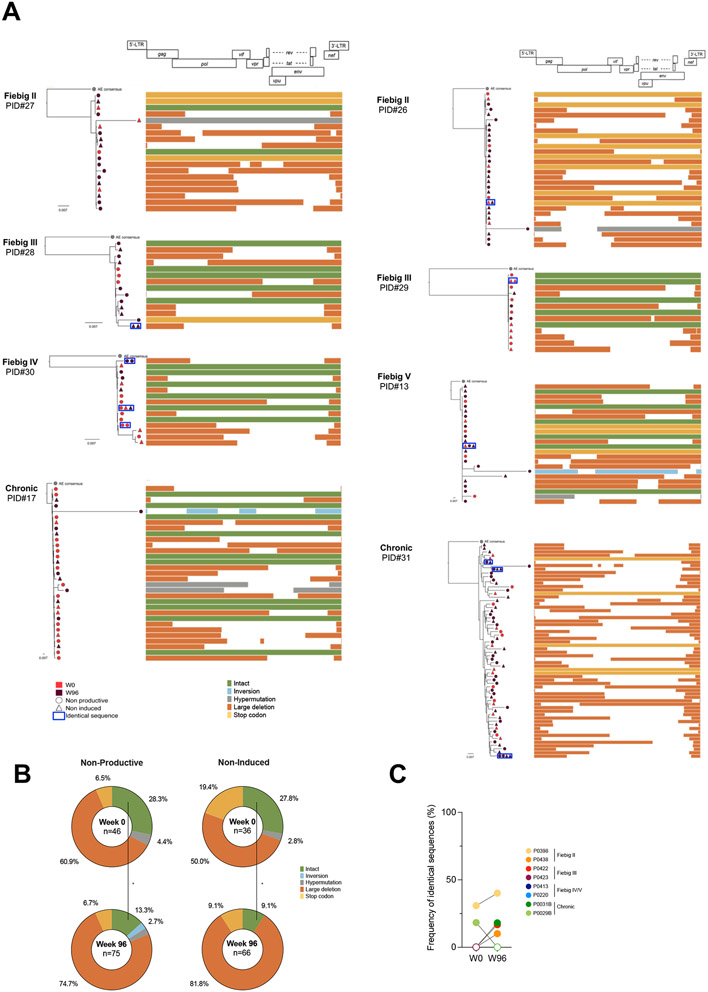
Upon infection, HIV disseminates throughout the human body within 1-2 weeks. However, its early cellular targets remain poorly characterized. We used a single-cell approach to retrieve the phenotype and TCR sequence of infected cells in blood and lymphoid tissue from individuals at the earliest stages of HIV infection. HIV initially targeted a few proliferating memory CD4+ T cells displaying high surface expression of CCR5. The phenotype of productively infected cells differed by Fiebig stage and between blood and lymph nodes. The TCR repertoire of productively infected cells was heavily biased, with preferential infection of previously expanded and disseminated clones, but composed almost exclusively of unique clonotypes, indicating that they were the product of independent infection events. Latent genetically intact proviruses were already archived early in infection. Hence, productive infection is initially established in a pool of phenotypically and clonotypically distinct T cells, and latently infected cells are generated simultaneously.
Keywords: HIV; HIV reservoir; acute infection; clonal expansion; inducibility; latency; lymph nodes; memory CD4(+) T cells; productive infection; proliferation.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Comment in
-
HIV establishes an early foothold.Cell Host Microbe. 2023 Apr 12;31(4):571-573. doi: 10.1016/j.chom.2023.03.012. Cell Host Microbe. 2023. PMID: 37054675
References
-
- Barre-Sinoussi F, Chermann JC, Rey F, Nugeyre MT, Chamaret S, Gruest J, Dauguet C, Axler-Blin C, Vezinet-Brun F, Rouzioux C, et al. (1983). Isolation of a T-lymphotropic retrovirus from a patient at risk for acquired immune deficiency syndrome (AIDS). Science 220, 868–871. 10.1126/science.6189183. - DOI - PubMed
-
- Zhang ZQ, Wietgrefe SW, Li Q, Shore MD, Duan L, Reilly C, Lifson JD, and Haase AT (2004). Roles of substrate availability and infection of resting and activated CD4+ T cells in transmission and acute simian immunodeficiency virus infection. Proc Natl Acad Sci U S A 101, 5640–5645. 10.1073/pnas.0308425101. - DOI - PMC - PubMed
-
- Gordon SN, Klatt NR, Bosinger SE, Brenchley JM, Milush JM, Engram JC, Dunham RM, Paiardini M, Klucking S, Danesh A, et al. (2007). Severe depletion of mucosal CD4+ T cells in AIDS-free simian immunodeficiency virus-infected sooty mangabeys. J Immunol 179, 3026–3034. 10.4049/jimmunol.179.5.3026. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
