

E3 ligase autoinhibition by C-degron mimicry maintains C-degron substrate fidelity
- PMID: 36805027
- PMCID: PMC10080726
- DOI: 10.1016/j.molcel.2023.01.019
E3 ligase autoinhibition by C-degron mimicry maintains C-degron substrate fidelity
Abstract
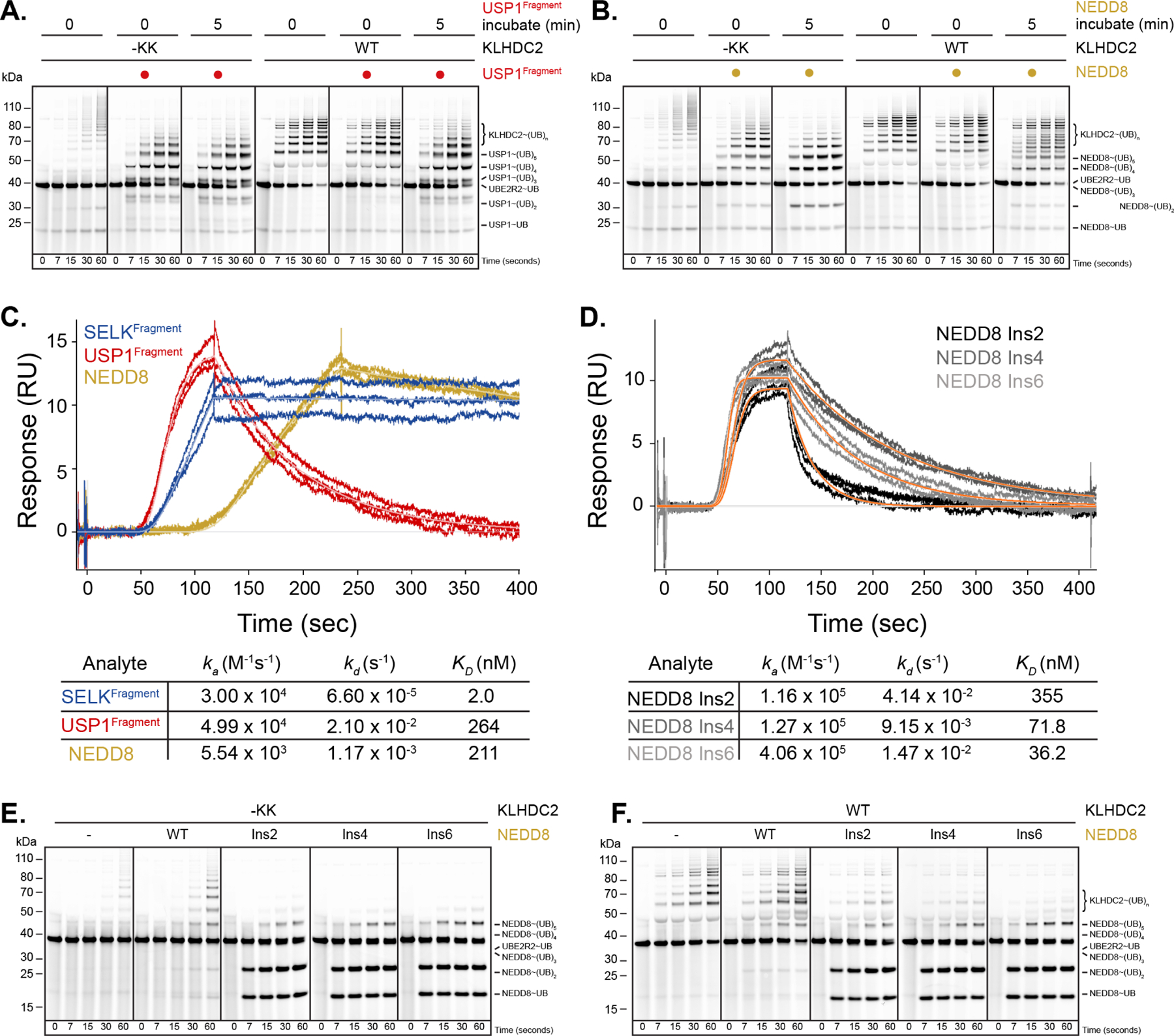
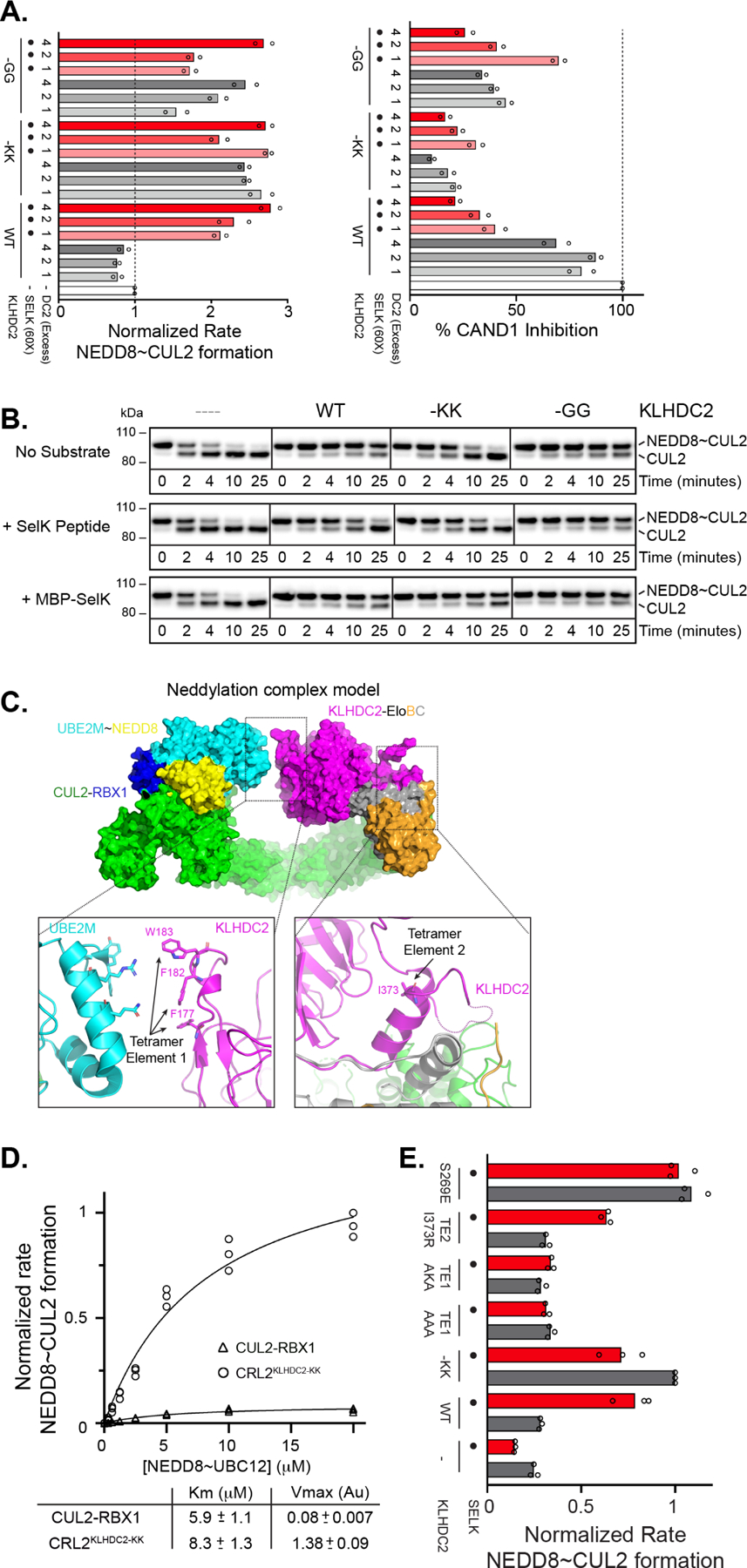
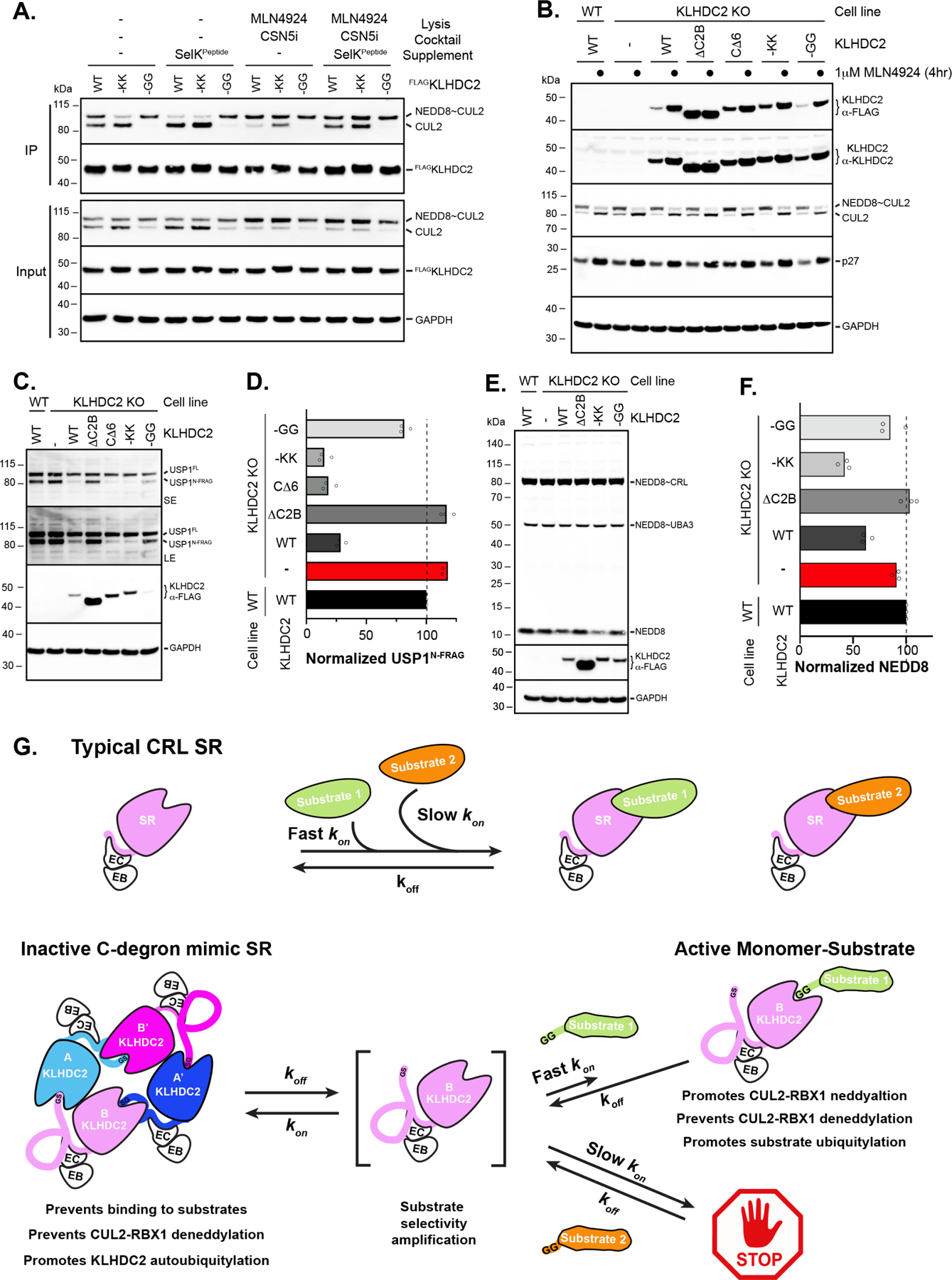
E3 ligase recruitment of proteins containing terminal destabilizing motifs (degrons) is emerging as a major form of regulation. How those E3s discriminate bona fide substrates from other proteins with terminal degron-like sequences remains unclear. Here, we report that human KLHDC2, a CRL2 substrate receptor targeting C-terminal Gly-Gly degrons, is regulated through interconversion between two assemblies. In the self-inactivated homotetramer, KLHDC2's C-terminal Gly-Ser motif mimics a degron and engages the substrate-binding domain of another protomer. True substrates capture the monomeric CRL2KLHDC2, driving E3 activation by neddylation and subsequent substrate ubiquitylation. Non-substrates such as NEDD8 bind KLHDC2 with high affinity, but its slow on rate prevents productive association with CRL2KLHDC2. Without substrate, neddylated CRL2KLHDC2 assemblies are deactivated via distinct mechanisms: the monomer by deneddylation and the tetramer by auto-ubiquitylation. Thus, substrate specificity is amplified by KLHDC2 self-assembly acting like a molecular timer, where only bona fide substrates may bind before E3 ligase inactivation.
Keywords: Autoinhibition; C-END degron; CUL2; Cullin-RING Ligase; E3; KLHDC10; KLHDC3; Kinetic proofreading; NEDD8; Targeted protein degradation; allostery; higher-order assembly; protein-protein interaction; ubiquitin.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests B.A.S. is a member of the scientific advisory boards of Interline Therapeutics and BioTheryX. B.A.S. and D.C.S. are co-inventors of intellectual property licensed to Cinsano.
Figures
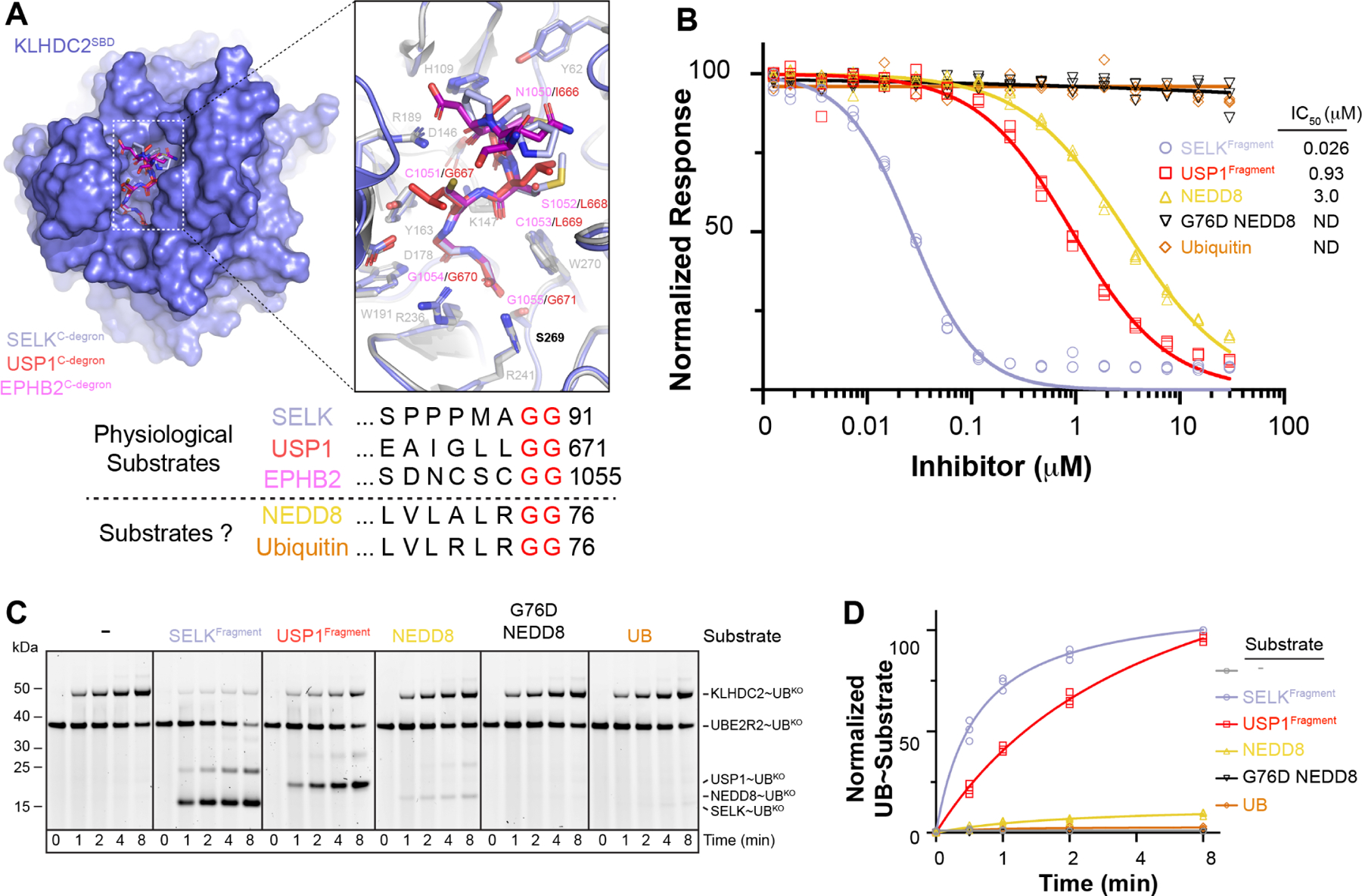
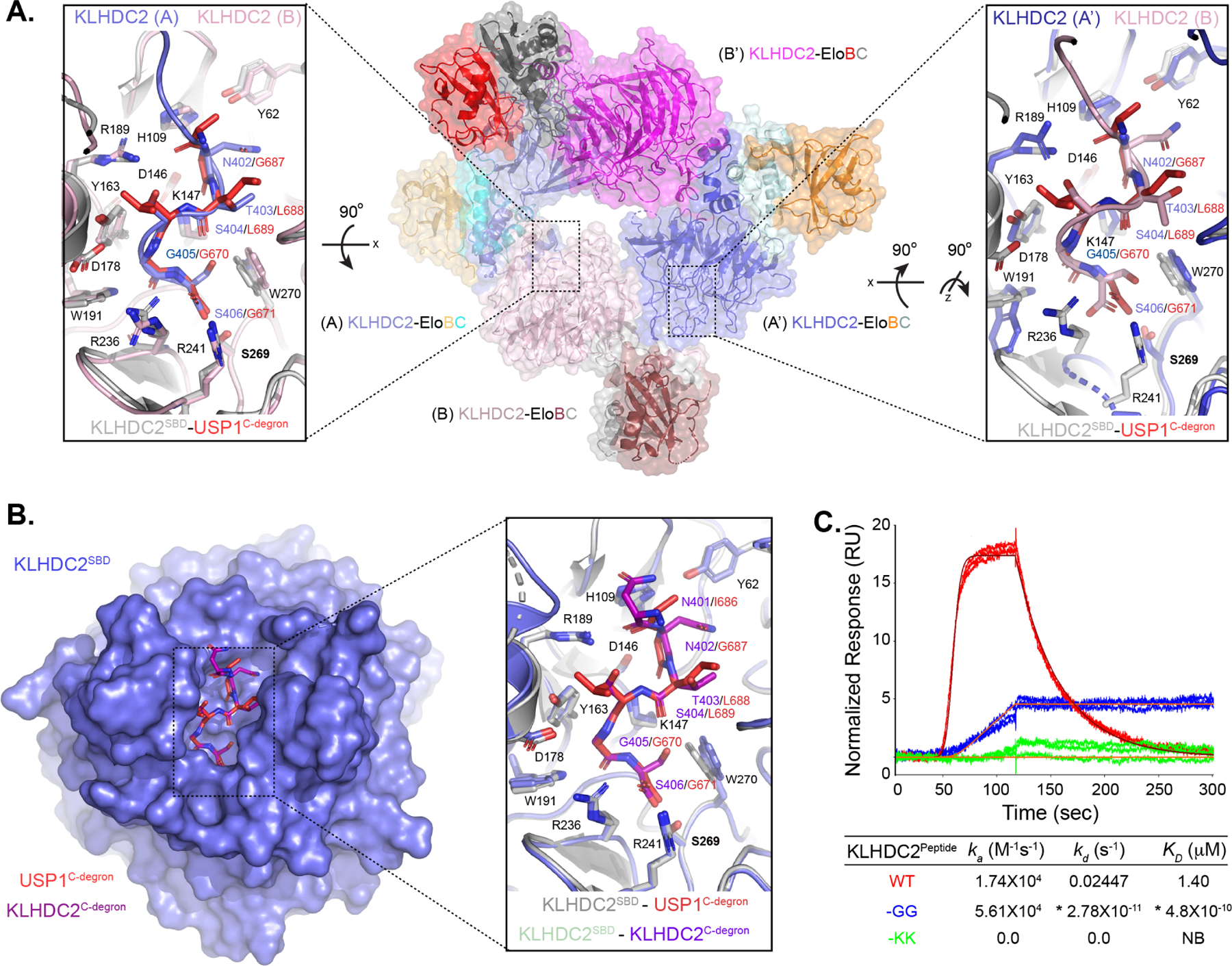
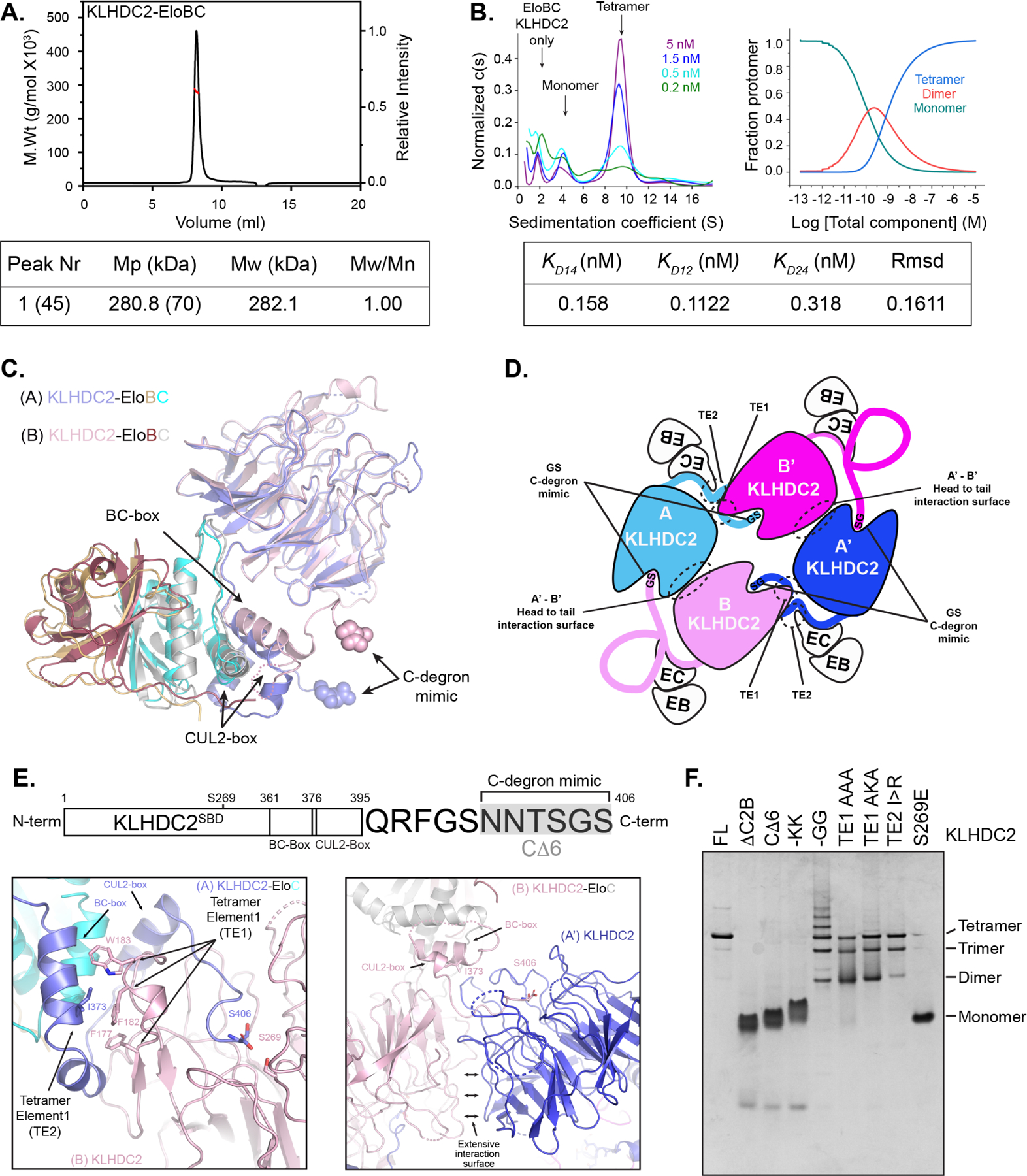
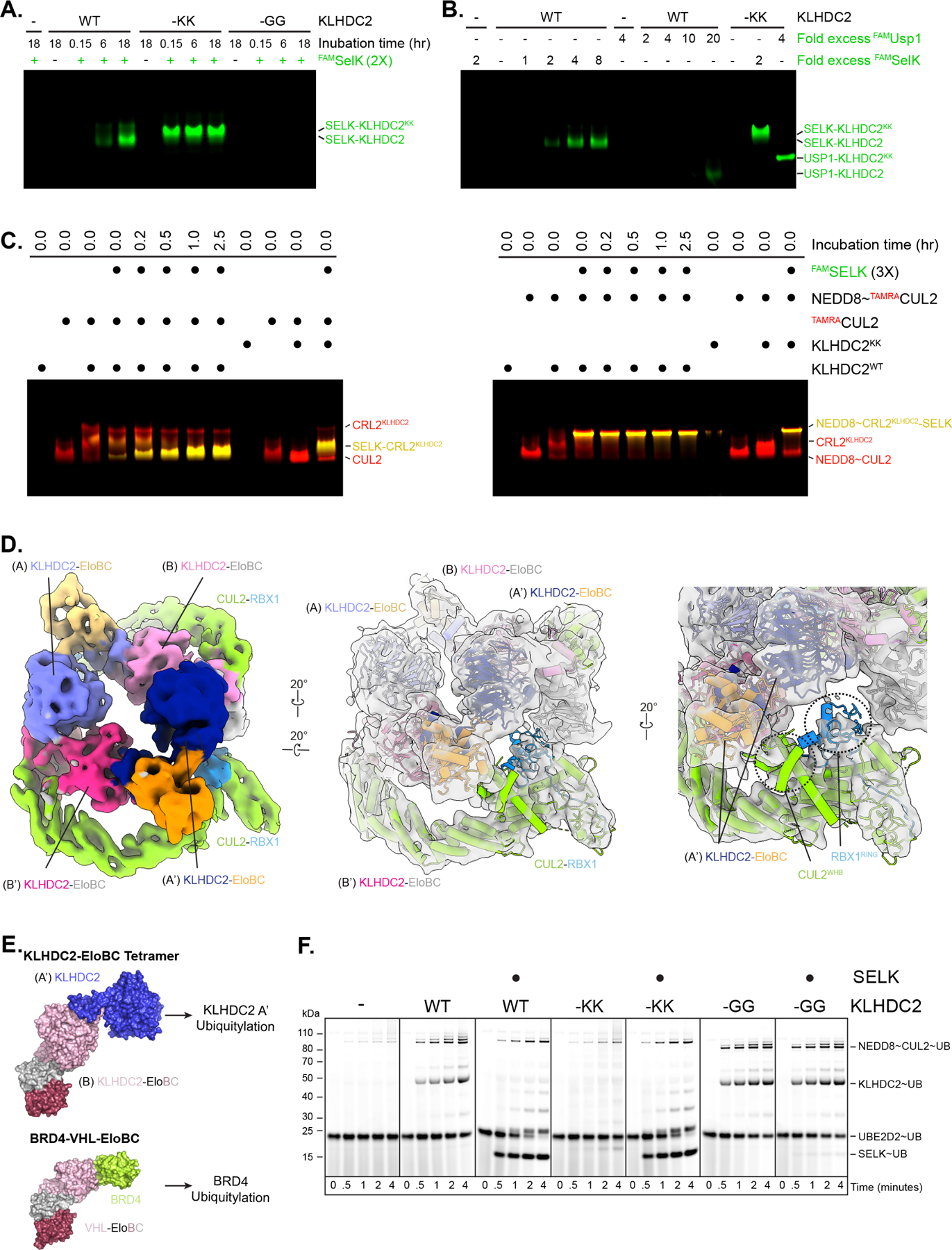
Comment in
-
C-degron mimicry confers E3 ligase selectivity.Nat Rev Mol Cell Biol. 2023 Apr;24(4):241. doi: 10.1038/s41580-023-00597-9. Nat Rev Mol Cell Biol. 2023. PMID: 36914761 No abstract available.
References
-
- Li W, Bengtson MH, Ulbrich A, Matsuda A, Reddy VA, Orth A, Chanda SK, Batalov S, and Joazeiro CA (2008). Genome-wide and functional annotation of human E3 ubiquitin ligases identifies MULAN, a mitochondrial E3 that regulates the organelle’s dynamics and signaling. PLoS One 3, e1487. 10.1371/journal.pone.0001487. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
