

Horizontal Gene Transfer and Fusion Spread Carotenogenesis Among Diverse Heterotrophic Protists
- PMID: 36805209
- PMCID: PMC10016063
- DOI: 10.1093/gbe/evad029
Horizontal Gene Transfer and Fusion Spread Carotenogenesis Among Diverse Heterotrophic Protists
Abstract
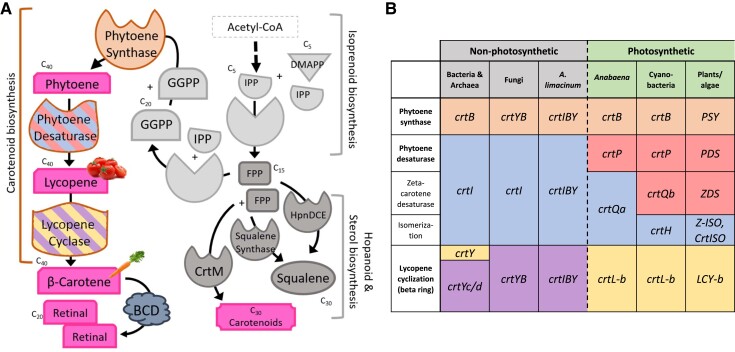
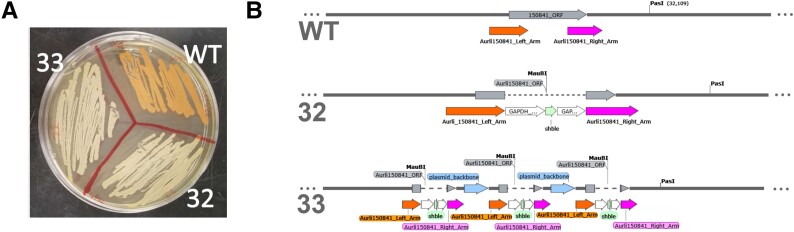
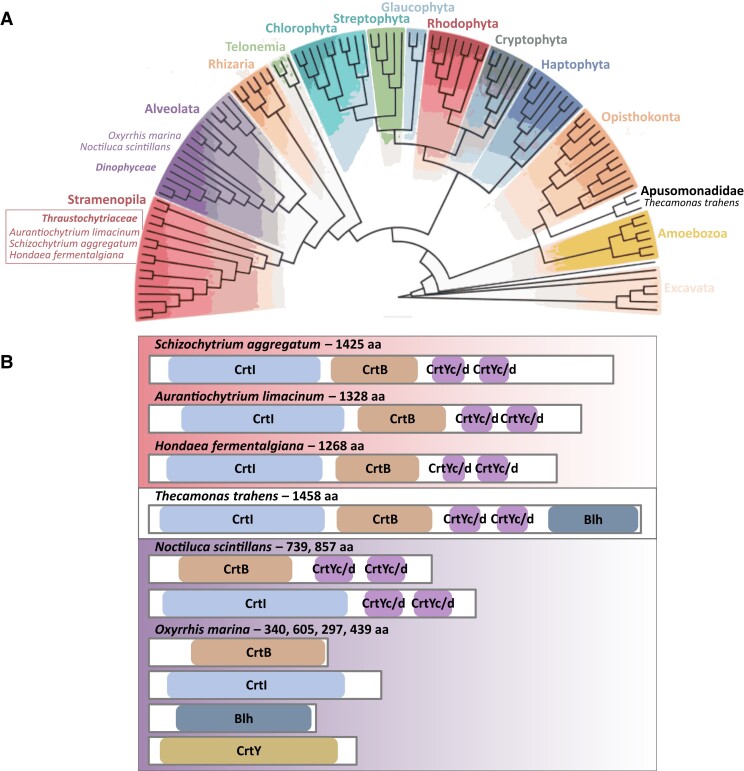
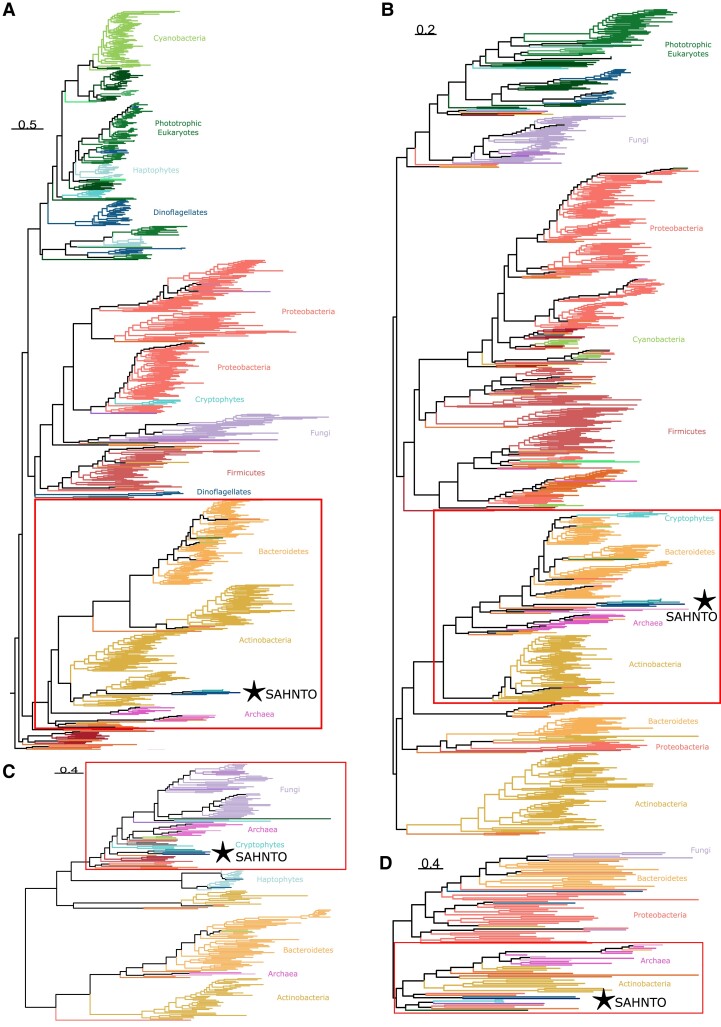
Thraustochytrids (phylum: Labyrinthulomycota) are nonphotosynthetic marine protists. Some thraustochytrids have crtIBY, a trifunctional fusion gene encoding a protein capable of β-carotene biosynthesis from geranylgeranyl pyrophosphate. Here we show that crtIBY is essential in, and encodes the sole pathway for, carotenoid biosynthesis in the thraustochytrid Aurantiochytrium limacinum ATCC MYA-1381. We explore the evolutionary origins of CrtIBY and discover that the closest related protein domains are present in a small but diverse group of other heterotrophic protists, including the apusomonad Thecamonas trahens and the dinoflagellates Oxyrrhis marina and Noctiluca scintillans. Each organism within this cluster also contains one or more β-carotene 15-15' oxygenase genes (blh and rpe65), suggesting that the acquisition of β-carotene biosynthesis genes may have been related to the production of retinal. Our findings support a novel origin of eukaryotic (apo)carotenoid biosynthesis by horizontal gene transfer from Actinobacteria, Bacteroidetes, and/or Archaea. This reveals a remarkable case of parallel evolution of eukaryotic (apo)carotenogenesis in divergent protistan lineages by repeated gene transfers.
Keywords: carotenoid oxygenase; lycopene cyclase; phylogenetics; phytoene desaturase; phytoene synthase; thraustochytrids.
© The Author(s) 2023. Published by Oxford University Press on behalf of Society for Molecular Biology and Evolution.
Figures

References
-
- Aki T, et al. . 2003. Thraustochytrid as a potential source of carotenoids. J Am Oil Chem Soc. 80:789–794.
-
- Alcaíno J, Baeza M, Cifuentes V. 2016. Carotenoid distribution in nature. In: Stange C, editors. Carotenoids in nature: biosynthesis, regulation and function. Vol. 79. Subcellular Biochemistry. Switzerland: Springer. p. 3–33. - PubMed
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. 1990. Basic local alignment search tool. J Mol Biol. 215(3):403–410. - PubMed
-
- Andrews AG, Starr MP. 1976. (3r,3'R)-Astaxanthin from the yeast Phaffia rhodozyma. Phytochemisty 15:1009–1011.
Publication types
MeSH terms
Substances
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
