

Enteroendocrine cell lineages that differentially control feeding and gut motility
- PMID: 36810133
- PMCID: PMC10032656
- DOI: 10.7554/eLife.78512
Enteroendocrine cell lineages that differentially control feeding and gut motility
Abstract
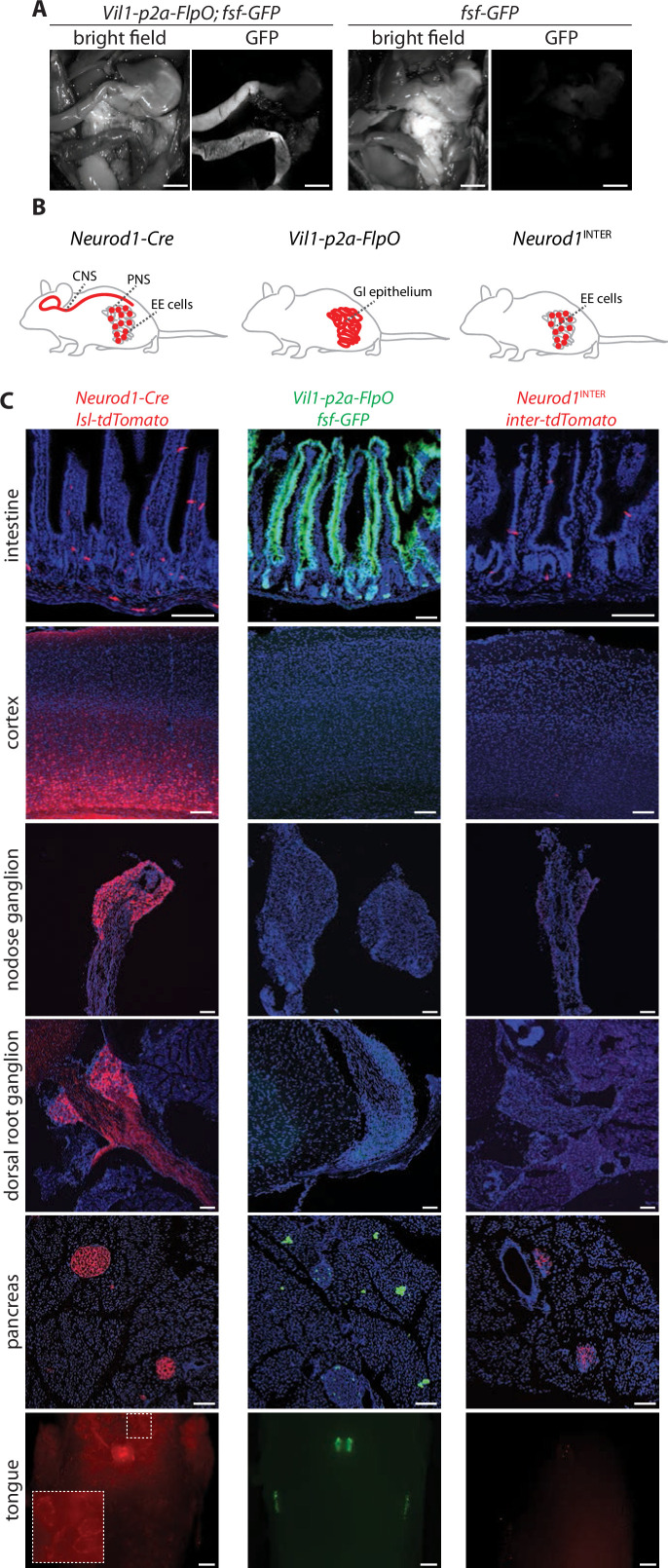
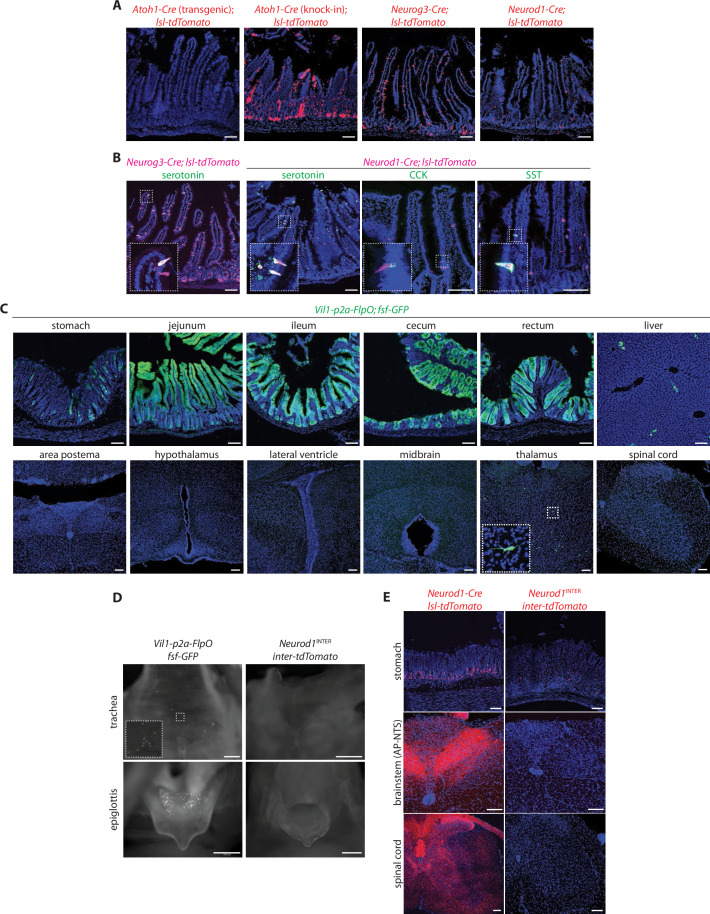
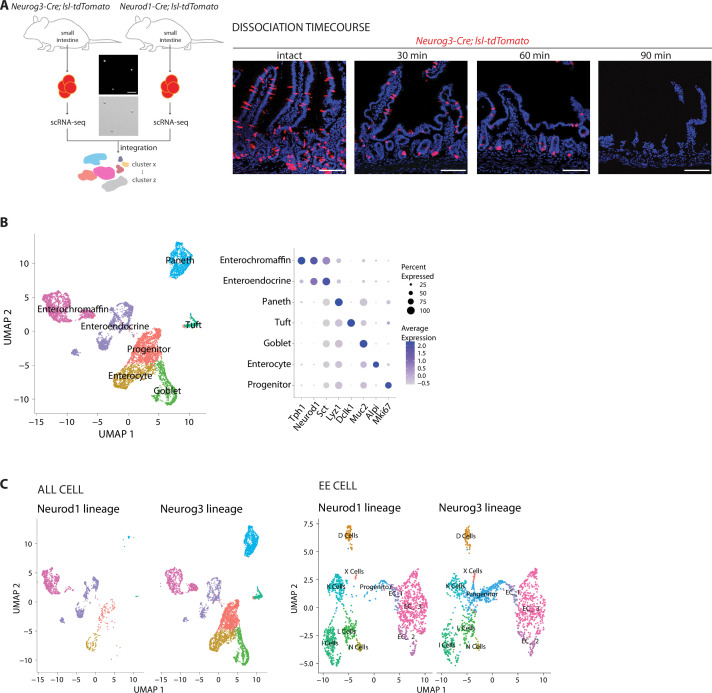
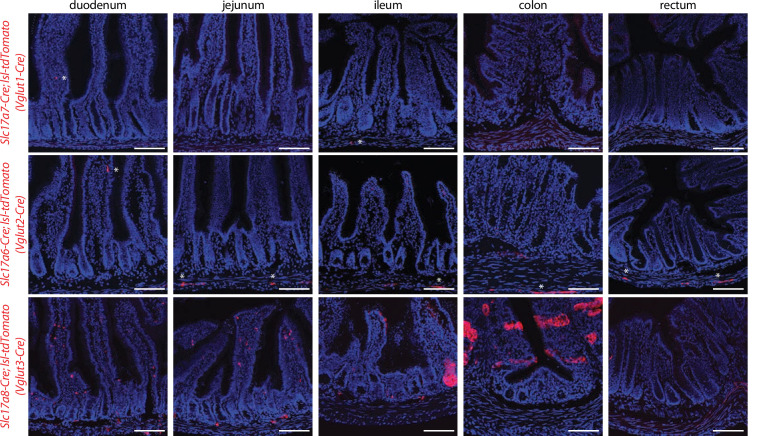
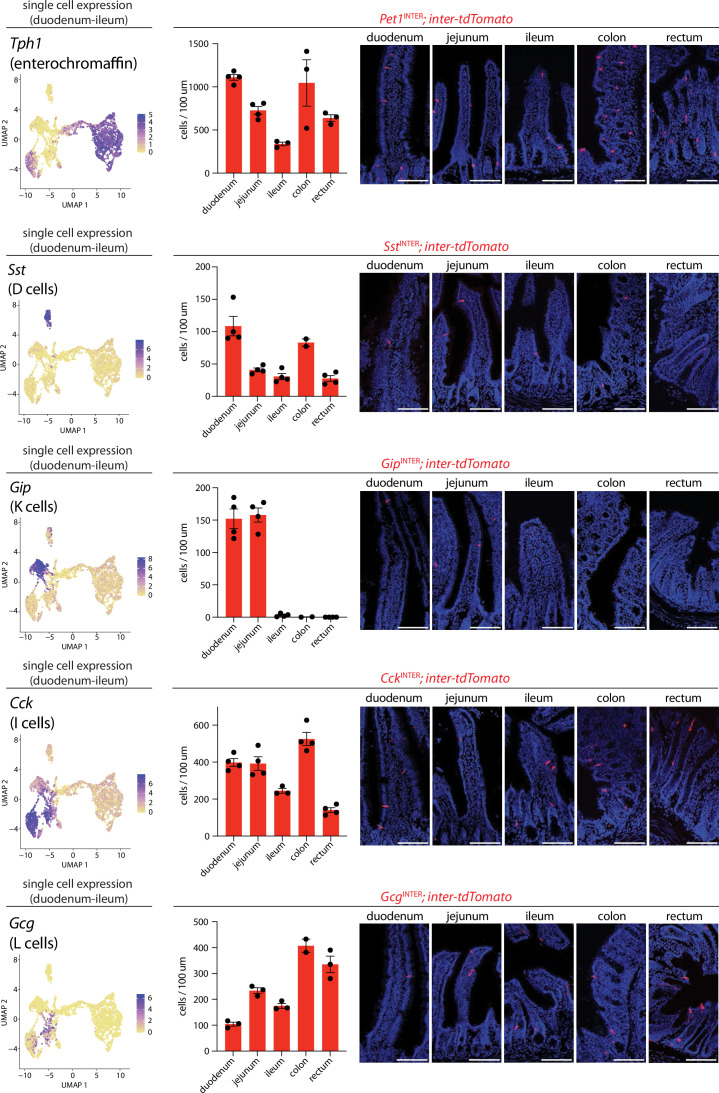
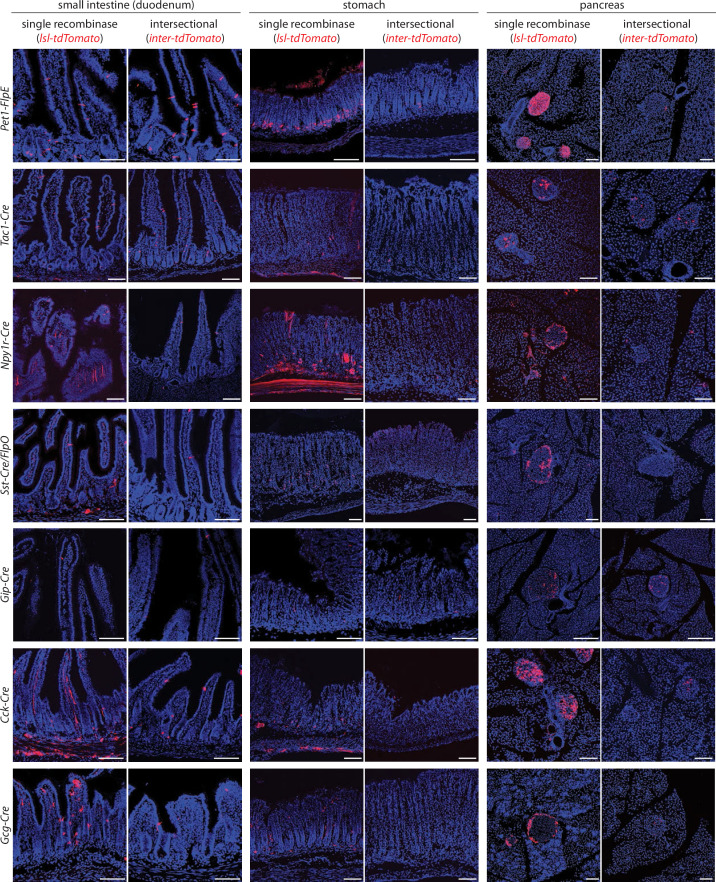
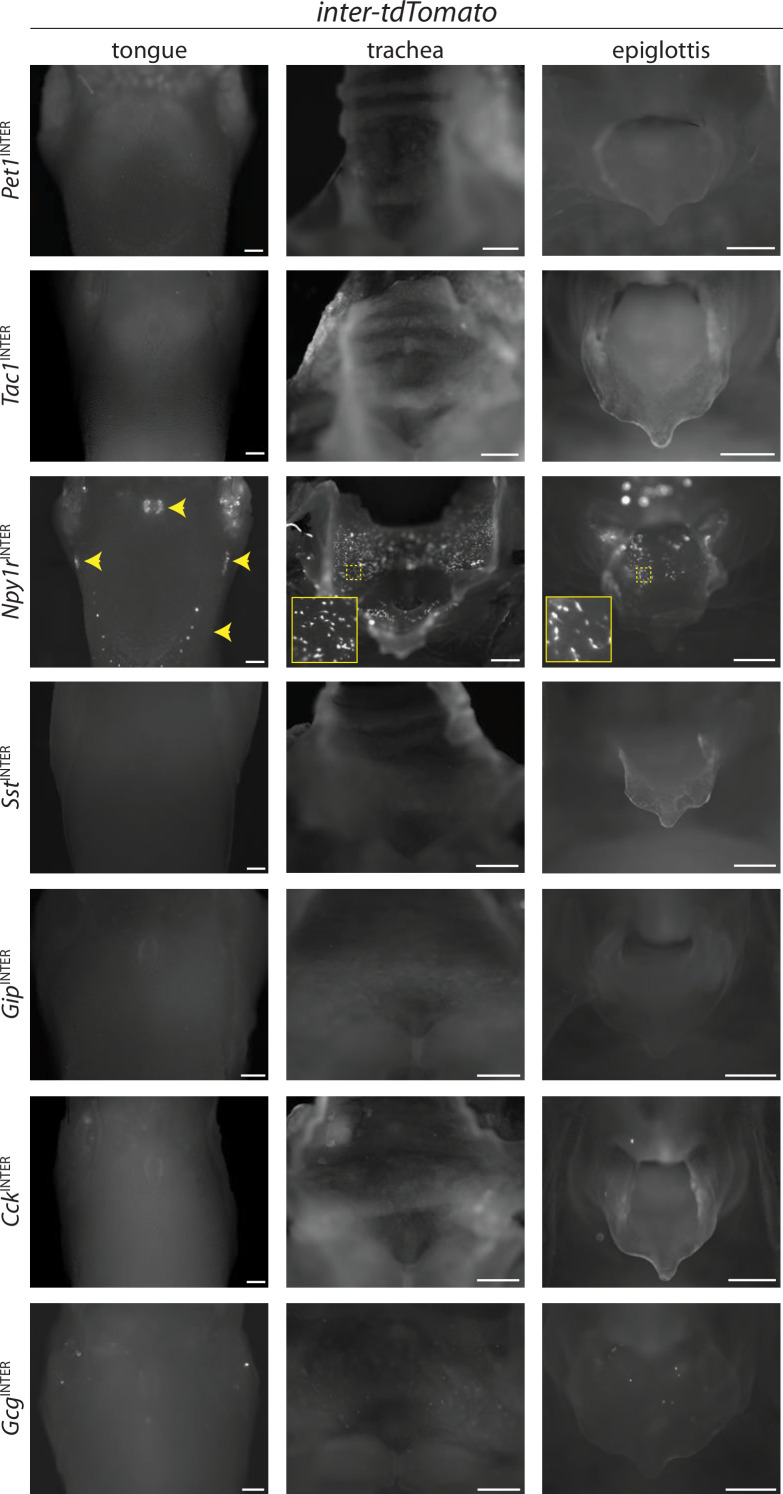
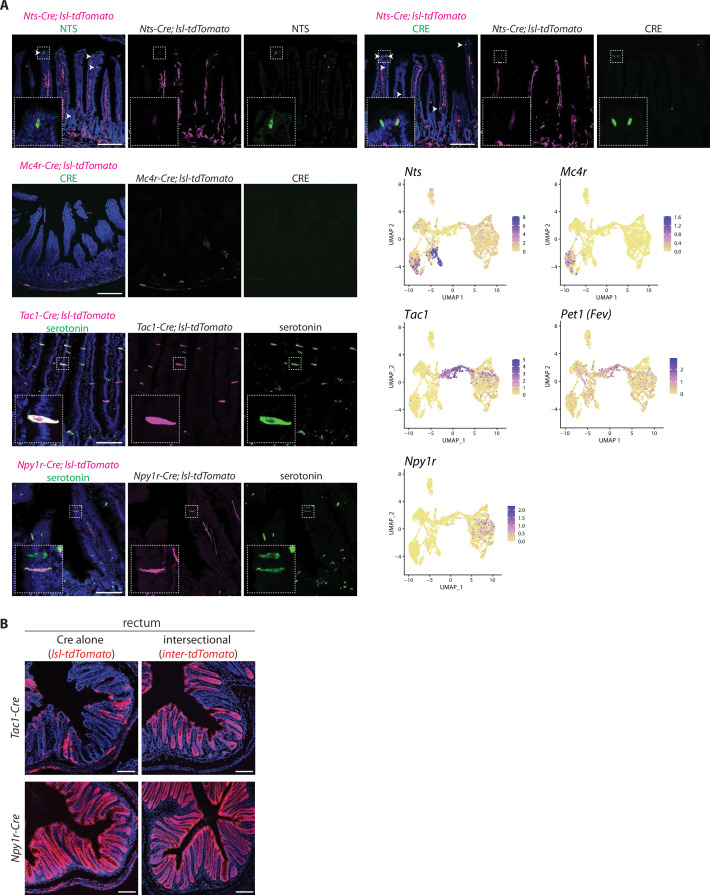
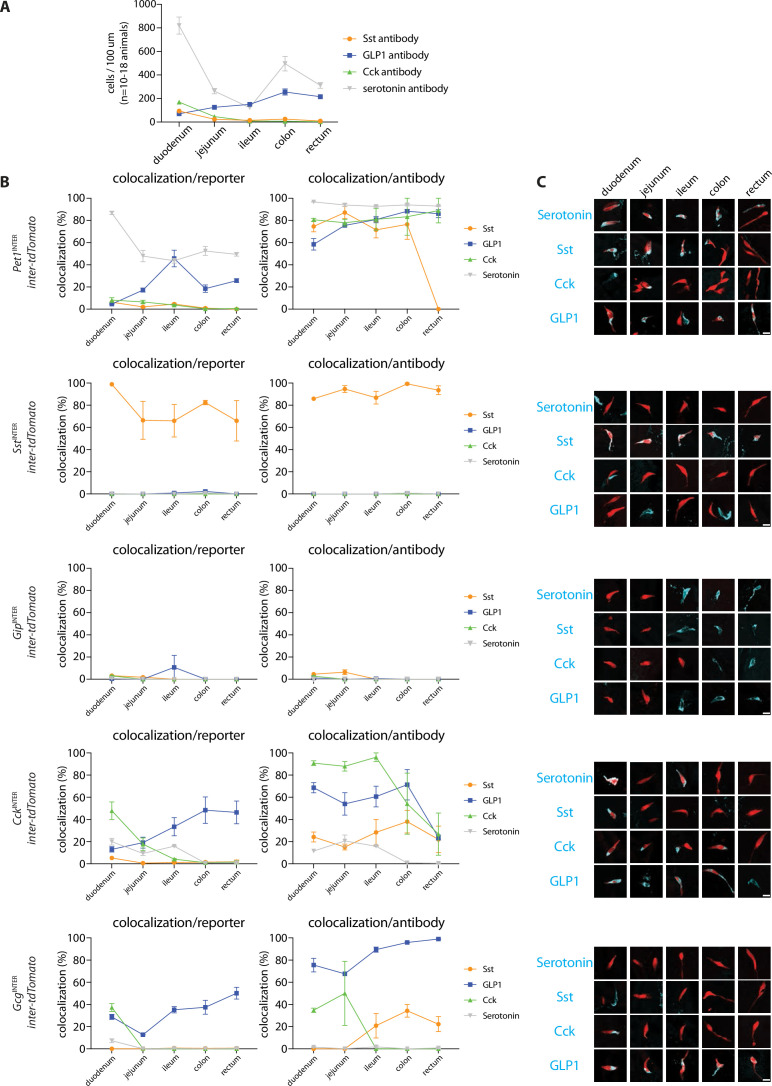
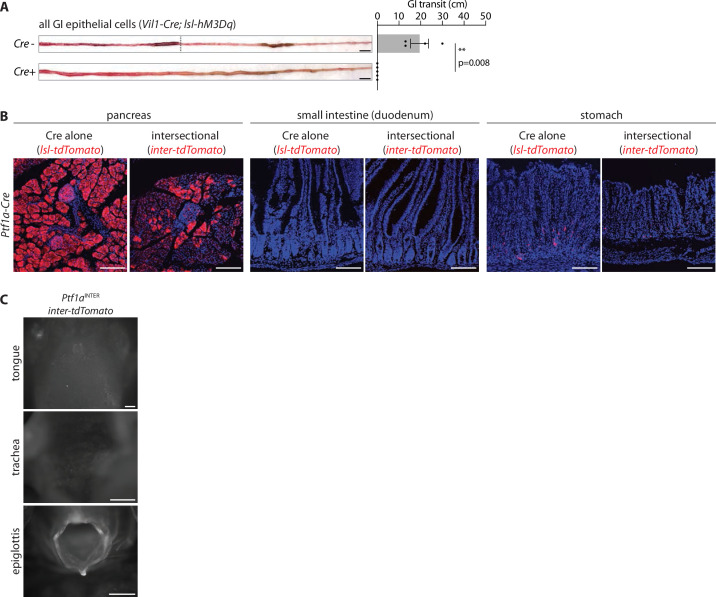
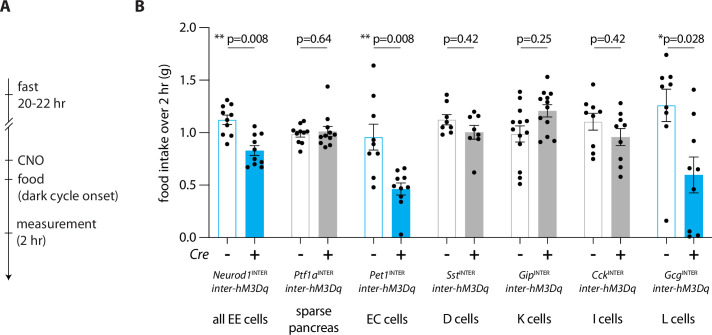
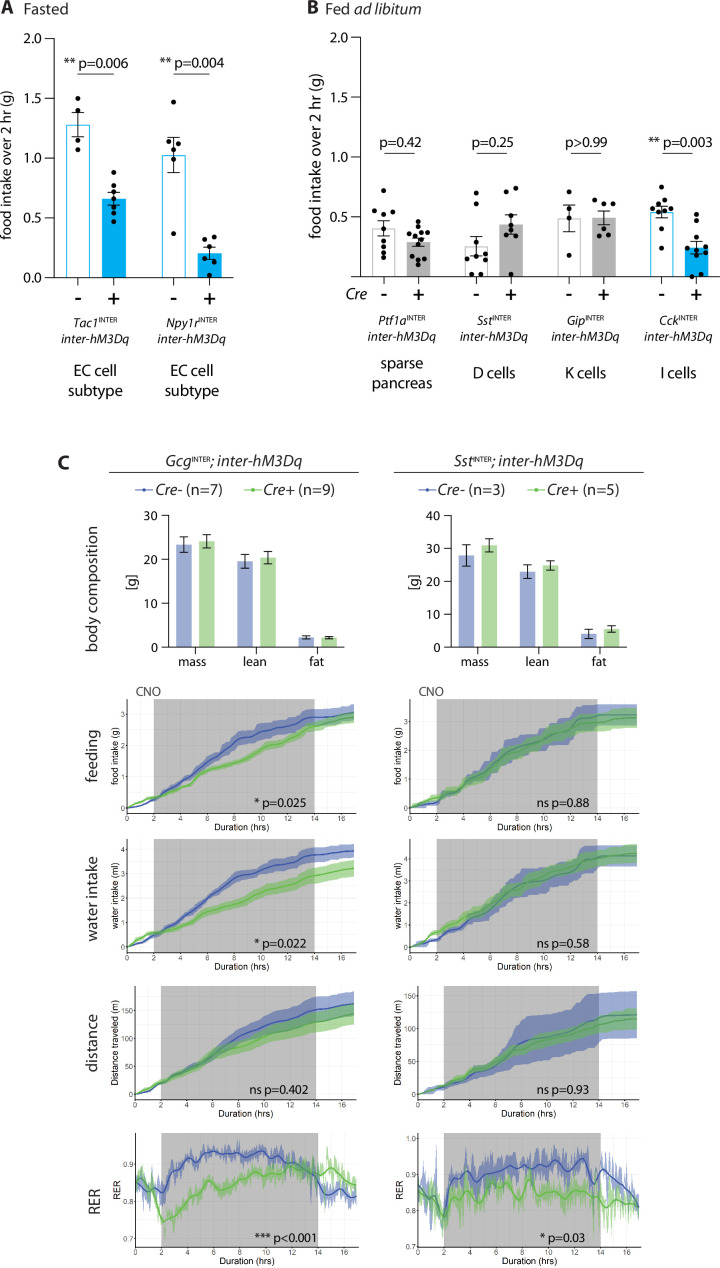
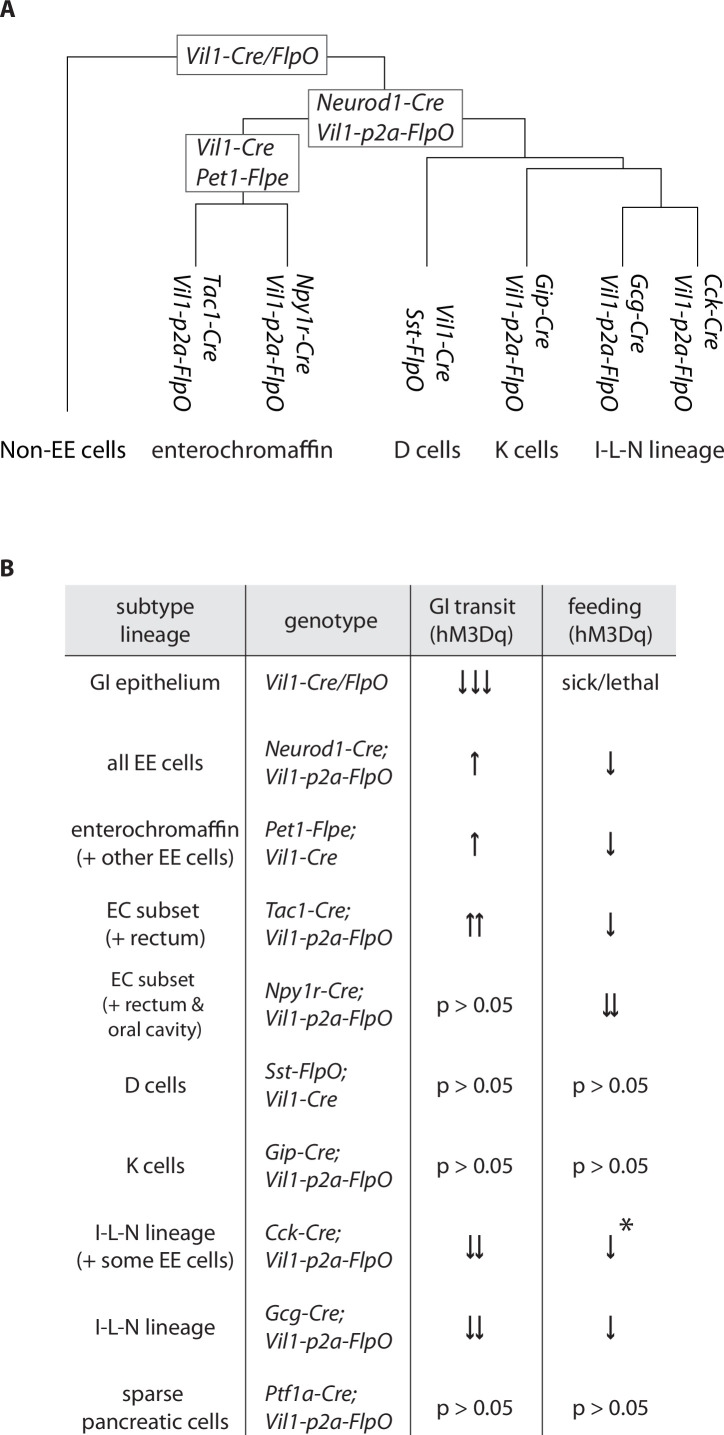
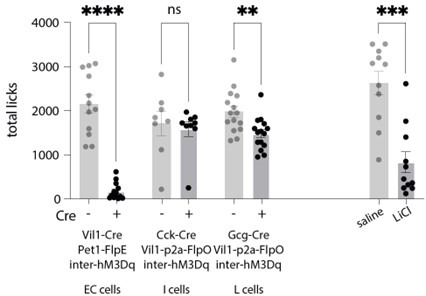
Enteroendocrine cells are specialized sensory cells of the gut-brain axis that are sparsely distributed along the intestinal epithelium. The functions of enteroendocrine cells have classically been inferred by the gut hormones they release. However, individual enteroendocrine cells typically produce multiple, sometimes apparently opposing, gut hormones in combination, and some gut hormones are also produced elsewhere in the body. Here, we developed approaches involving intersectional genetics to enable selective access to enteroendocrine cells in vivo in mice. We targeted FlpO expression to the endogenous Villin1 locus (in Vil1-p2a-FlpO knock-in mice) to restrict reporter expression to intestinal epithelium. Combined use of Cre and Flp alleles effectively targeted major transcriptome-defined enteroendocrine cell lineages that produce serotonin, glucagon-like peptide 1, cholecystokinin, somatostatin, or glucose-dependent insulinotropic polypeptide. Chemogenetic activation of different enteroendocrine cell types variably impacted feeding behavior and gut motility. Defining the physiological roles of different enteroendocrine cell types provides an essential framework for understanding sensory biology of the intestine.
Keywords: appetite; enteroendocrine cell; gut-brain axis; intersectional genetics; mouse; neuroscience; single-cell RNA sequencing.
© 2023, Hayashi et al.
Conflict of interest statement
MH, JK, ED, NJ, FR No competing interests declared, FG Is a consultant for Kallyope, Inc, SL Reviewing editor, eLife
Figures


Update of
References
-
- Alcaino C, Knutson KR, Treichel AJ, Yildiz G, Strege PR, Linden DR, Li JH, Leiter AB, Szurszewski JH, Farrugia G, Beyder A. A population of gut epithelial enterochromaffin cells is mechanosensitive and requires piezo2 to convert force into serotonin release. PNAS. 2018;115:E7632–E7641. doi: 10.1073/pnas.1804938115. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
