

Involvement of GABAergic Interneuron Subtypes in 4-Aminopyridine-Induced Seizure-Like Events in Mouse Entorhinal Cortex in Vitro
- PMID: 36810229
- PMCID: PMC10027059
- DOI: 10.1523/JNEUROSCI.1190-22.2023
Involvement of GABAergic Interneuron Subtypes in 4-Aminopyridine-Induced Seizure-Like Events in Mouse Entorhinal Cortex in Vitro
Abstract
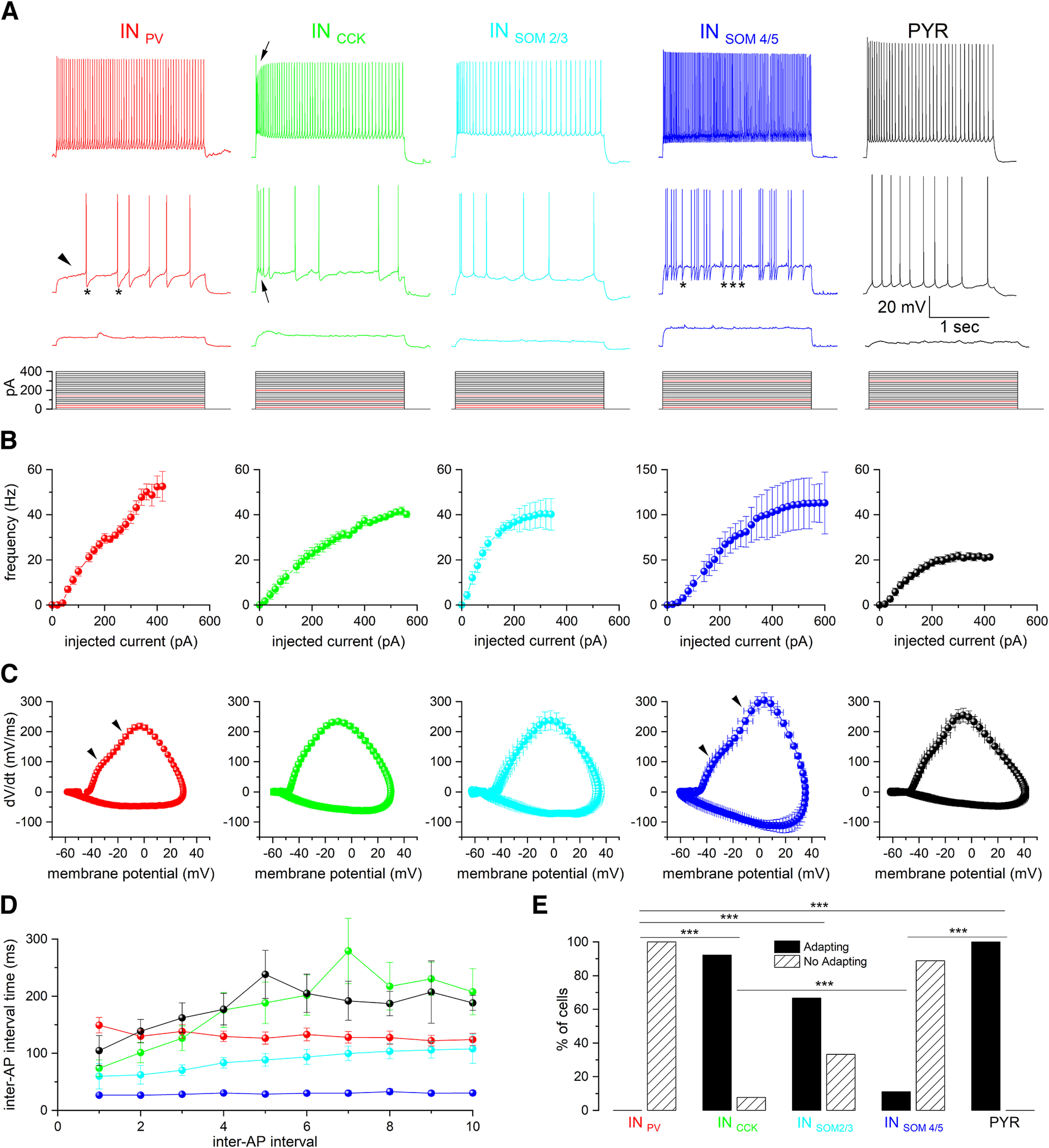
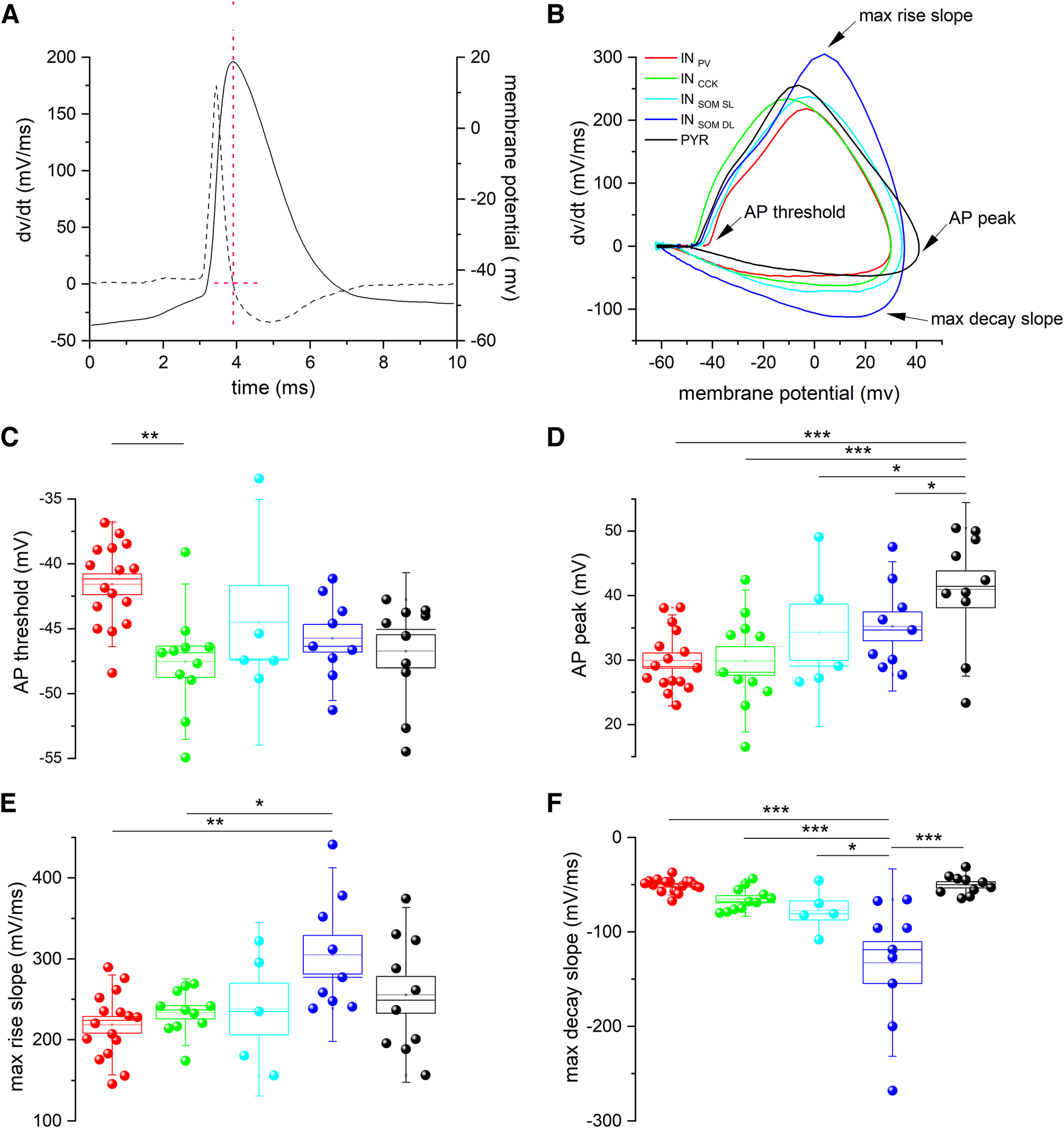
Single-unit recordings performed in temporal lobe epilepsy patients and in models of temporal lobe seizures have shown that interneurons are active at focal seizure onset. We performed simultaneous patch-clamp and field potential recordings in entorhinal cortex slices of GAD65 and GAD67 C57BL/6J male mice that express green fluorescent protein in GABAergic neurons to analyze the activity of specific interneuron (IN) subpopulations during acute seizure-like events (SLEs) induced by 4-aminopyridine (4-AP; 100 μm). IN subtypes were identified as parvalbuminergic (INPV, n = 17), cholecystokinergic (INCCK), n = 13], and somatostatinergic (INSOM, n = 15), according to neurophysiological features and single-cell digital PCR. INPV and INCCK discharged at the start of 4-AP-induced SLEs characterized by either low-voltage fast or hyper-synchronous onset pattern. In both SLE onset types, INSOM fired earliest before SLEs, followed by INPV and INCCK discharges. Pyramidal neurons became active with variable delays after SLE onset. Depolarizing block was observed in ∼50% of cells in each INs subgroup, and it was longer in IN (∼4 s) than in pyramidal neurons (<1 s). As SLE evolved, all IN subtypes generated action potential bursts synchronous with the field potential events leading to SLE termination. High-frequency firing throughout the SLE occurred in one-third of INPV and INSOM We conclude that entorhinal cortex INs are very active at the onset and during the progression of SLEs induced by 4-AP. These results support earlier in vivo and in vivo evidence and suggest that INs have a preferential role in focal seizure initiation and development.SIGNIFICANCE STATEMENT Focal seizures are believed to result from enhanced excitation. Nevertheless, we and others demonstrated that cortical GABAergic networks may initiate focal seizures. Here, we analyzed for the first time the role of different IN subtypes in seizures generated by 4-aminopyridine in the mouse entorhinal cortex slices. We found that in this in vitro focal seizure model, all IN types contribute to seizure initiation and that INs precede firing of principal cells. This evidence is in agreement with the active role of GABAergic networks in seizure generation.
Keywords: epilepsy; in vitro slices; interneurons; patch clamp; seizures.
Copyright © 2023 the authors.
Figures







Similar articles
-
Interneuronal Network Activity at the Onset of Seizure-Like Events in Entorhinal Cortex Slices.J Neurosci. 2017 Oct 25;37(43):10398-10407. doi: 10.1523/JNEUROSCI.3906-16.2017. Epub 2017 Sep 25. J Neurosci. 2017. PMID: 28947576 Free PMC article.
-
Interneuron-Driven Ictogenesis in the 4-Aminopyridine Model: Depolarization Block and Potassium Accumulation Initiate Seizure-like Activity.Int J Mol Sci. 2025 Jul 16;26(14):6812. doi: 10.3390/ijms26146812. Int J Mol Sci. 2025. PMID: 40725059 Free PMC article.
-
Synchronous inhibitory potentials precede seizure-like events in acute models of focal limbic seizures.J Neurosci. 2015 Feb 18;35(7):3048-55. doi: 10.1523/JNEUROSCI.3692-14.2015. J Neurosci. 2015. PMID: 25698742 Free PMC article.
-
GABAA-Receptor Signaling and Ionic Plasticity in the Generation and Spread of Seizures.In: Noebels JL, Avoli M, Rogawski MA, Vezzani A, Delgado-Escueta AV, editors. Jasper's Basic Mechanisms of the Epilepsies. 5th edition. New York: Oxford University Press; 2024. Chapter 6. In: Noebels JL, Avoli M, Rogawski MA, Vezzani A, Delgado-Escueta AV, editors. Jasper's Basic Mechanisms of the Epilepsies. 5th edition. New York: Oxford University Press; 2024. Chapter 6. PMID: 39637123 Free Books & Documents. Review.
-
Mechanisms Leading to Initiation, Development, and Termination of Focal Seizures.In: Noebels JL, Avoli M, Rogawski MA, Vezzani A, Delgado-Escueta AV, editors. Jasper's Basic Mechanisms of the Epilepsies. 5th edition. New York: Oxford University Press; 2024. Chapter 8. In: Noebels JL, Avoli M, Rogawski MA, Vezzani A, Delgado-Escueta AV, editors. Jasper's Basic Mechanisms of the Epilepsies. 5th edition. New York: Oxford University Press; 2024. Chapter 8. PMID: 39637112 Free Books & Documents. Review.
Cited by
-
Elevated susceptibility to exogenous seizure triggers and impaired interneuron excitability in a mouse model of Leigh syndrome epilepsy.Neurobiol Dis. 2023 Oct 15;187:106288. doi: 10.1016/j.nbd.2023.106288. Epub 2023 Sep 11. Neurobiol Dis. 2023. PMID: 37704057 Free PMC article.
-
Cracking the code: the clinical and molecular impact of aminopyridines; a review (2019-2024).RSC Adv. 2025 Jan 8;15(1):688-711. doi: 10.1039/d4ra07438f. eCollection 2025 Jan 2. RSC Adv. 2025. PMID: 39781020 Free PMC article. Review.
-
Fast activity chirp patterns in focal seizures from patients and animal models.Epilepsia. 2025 Mar;66(3):621-631. doi: 10.1111/epi.18245. Epub 2024 Dec 26. Epilepsia. 2025. PMID: 39723840 Free PMC article. Review.
-
Modulation of in vitro Network Activity by Optogenetic Stimulation of Parvalbumin-positive Interneurons During Estrous Cycle.Curr Neuropharmacol. 2025;23(7):862-871. doi: 10.2174/011570159X326861241129093354. Curr Neuropharmacol. 2025. PMID: 39844403 Free PMC article.
-
Downmodulation of Potassium Conductances Induces Epileptic Seizures in Cortical Network Models Via Multiple Synergistic Factors.J Neurosci. 2025 Mar 26;45(13):e1909232025. doi: 10.1523/JNEUROSCI.1909-23.2025. J Neurosci. 2025. PMID: 39880680
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials