

Mutant structure of metabolic switch protein in complex with monomeric c-di-GMP reveals a potential mechanism of protein-mediated ligand dimerization
- PMID: 36810577
- PMCID: PMC9944927
- DOI: 10.1038/s41598-023-29110-0
Mutant structure of metabolic switch protein in complex with monomeric c-di-GMP reveals a potential mechanism of protein-mediated ligand dimerization
Abstract
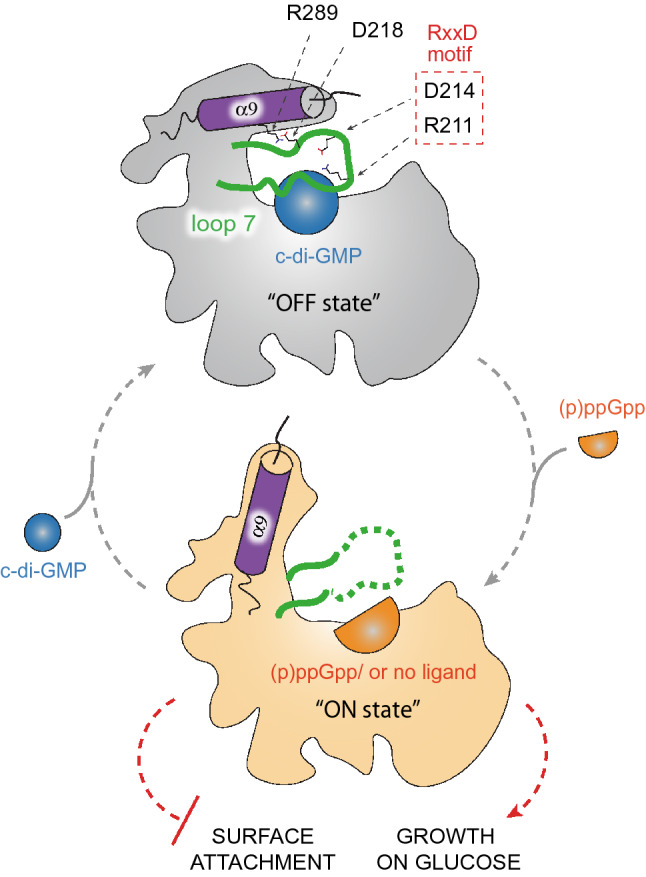
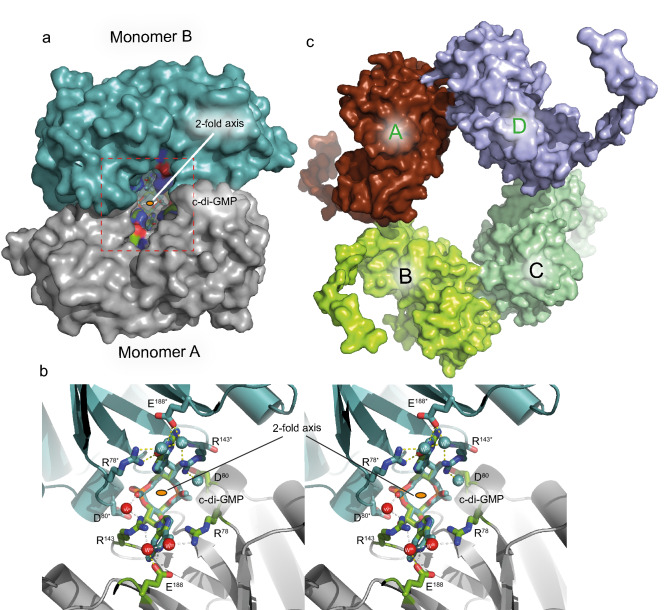
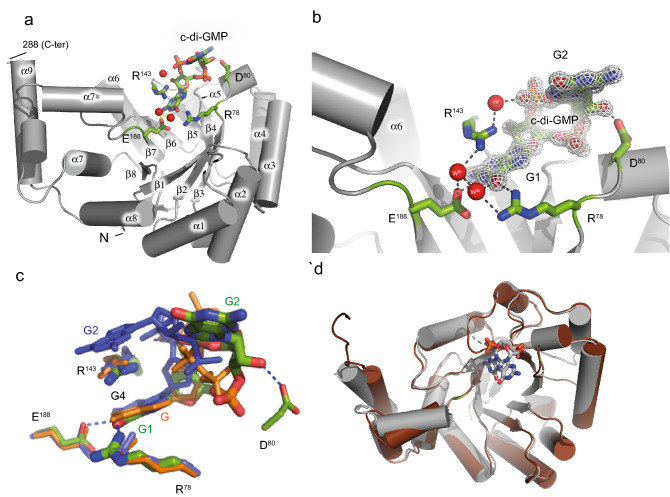
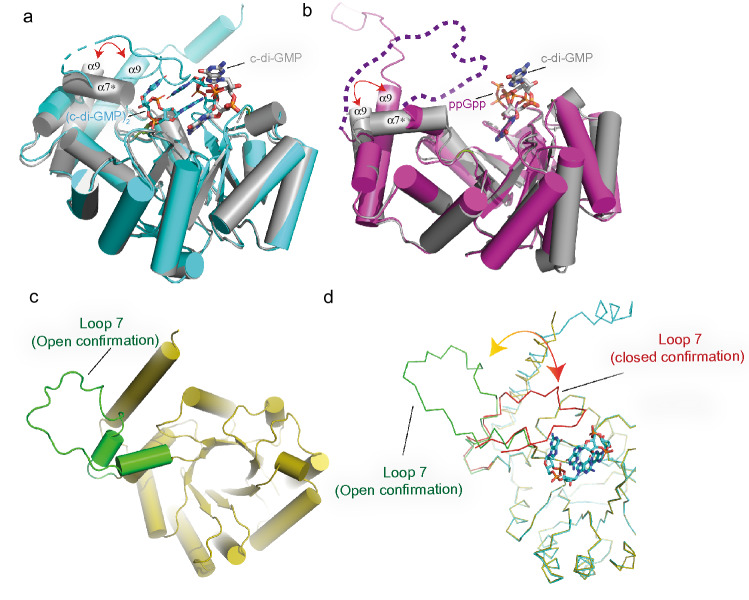
Bacterial second messengers c-di-GMP and (p)ppGpp have broad functional repertoires ranging from growth and cell cycle control to the regulation of biofilm formation and virulence. The recent identification of SmbA, an effector protein from Caulobacter crescentus that is jointly targeted by both signaling molecules, has opened up studies on how these global bacterial networks interact. C-di-GMP and (p)ppGpp compete for the same SmbA binding site, with a dimer of c-di-GMP inducing a conformational change that involves loop 7 of the protein that leads to downstream signaling. Here, we report a crystal structure of a partial loop 7 deletion mutant, SmbA∆loop in complex with c-di-GMP determined at 1.4 Å resolution. SmbA∆loop binds monomeric c-di-GMP indicating that loop 7 is required for c-di-GMP dimerization. Thus the complex probably represents the first step of consecutive c-di-GMP binding to form an intercalated dimer as has been observed in wild-type SmbA. Considering the prevalence of intercalated c-di-GMP molecules observed bound to proteins, the proposed mechanism may be generally applicable to protein-mediated c-di-GMP dimerization. Notably, in the crystal, SmbA∆loop forms a 2-fold symmetric dimer via isologous interactions with the two symmetric halves of c-di-GMP. Structural comparisons of SmbA∆loop with wild-type SmbA in complex with dimeric c-di-GMP or ppGpp support the idea that loop 7 is critical for SmbA function by interacting with downstream partners. Our results also underscore the flexibility of c-di-GMP, to allow binding to the symmetric SmbA∆loop dimer interface. It is envisaged that such isologous interactions of c-di-GMP could be observed in hitherto unrecognized targets.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
