

circFNDC3B Accelerates Vasculature Formation and Metastasis in Oral Squamous Cell Carcinoma
- PMID: 36811957
- PMCID: PMC10152237
- DOI: 10.1158/0008-5472.CAN-22-2585
circFNDC3B Accelerates Vasculature Formation and Metastasis in Oral Squamous Cell Carcinoma
Abstract
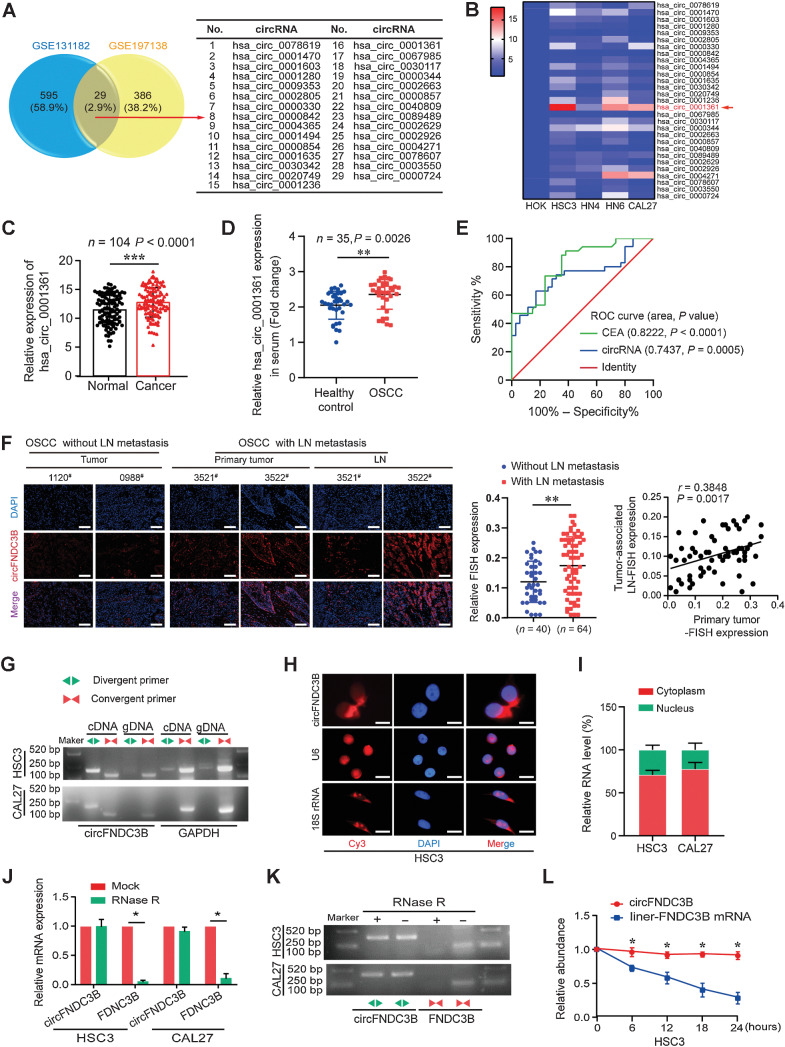
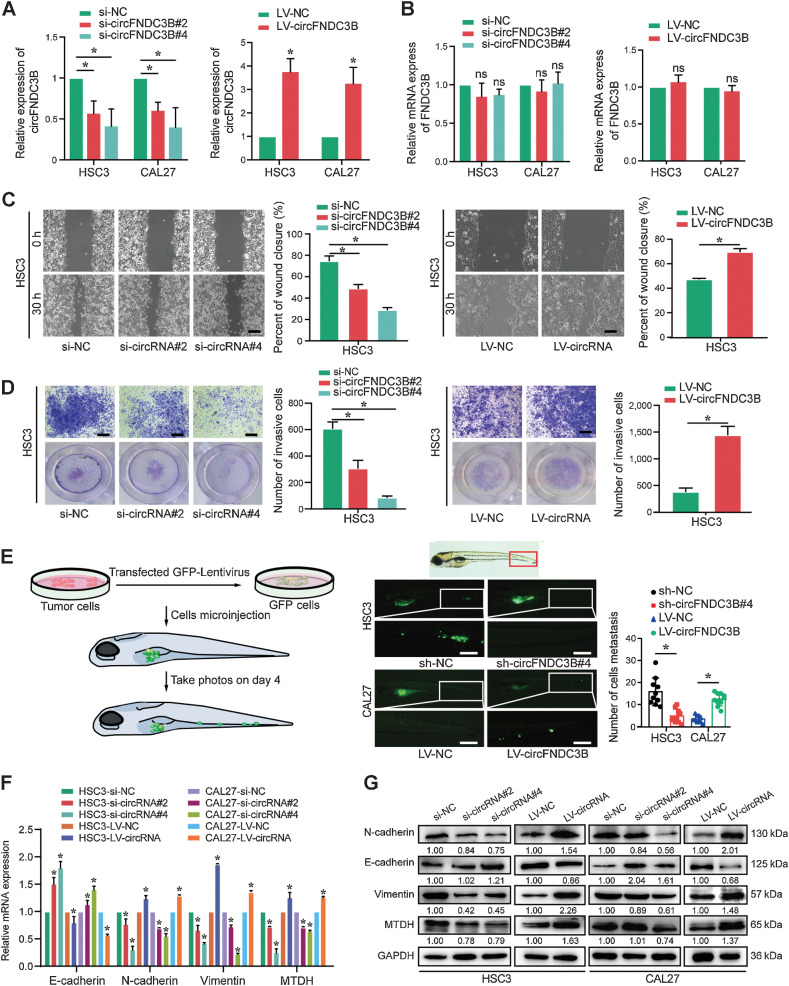
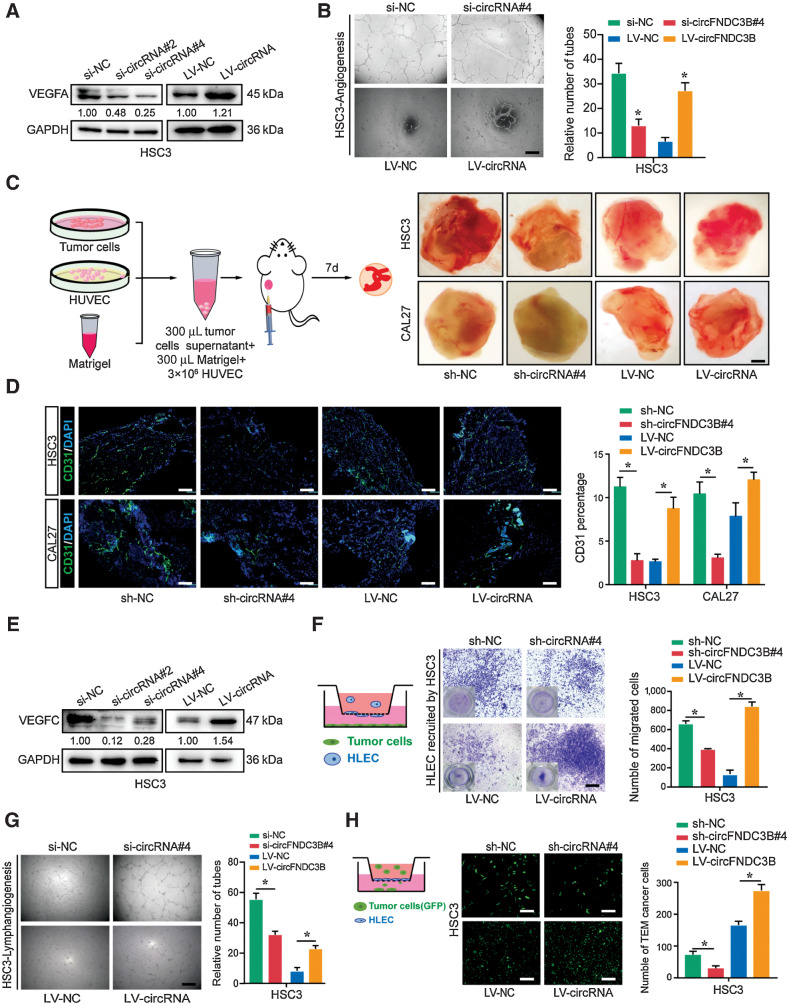
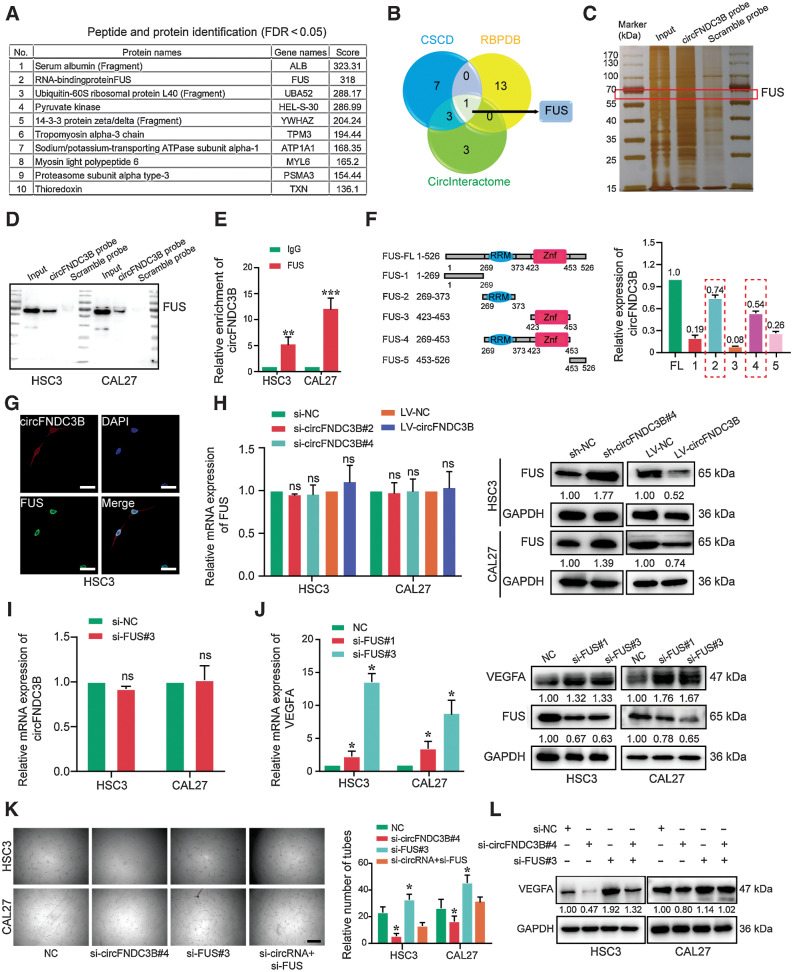
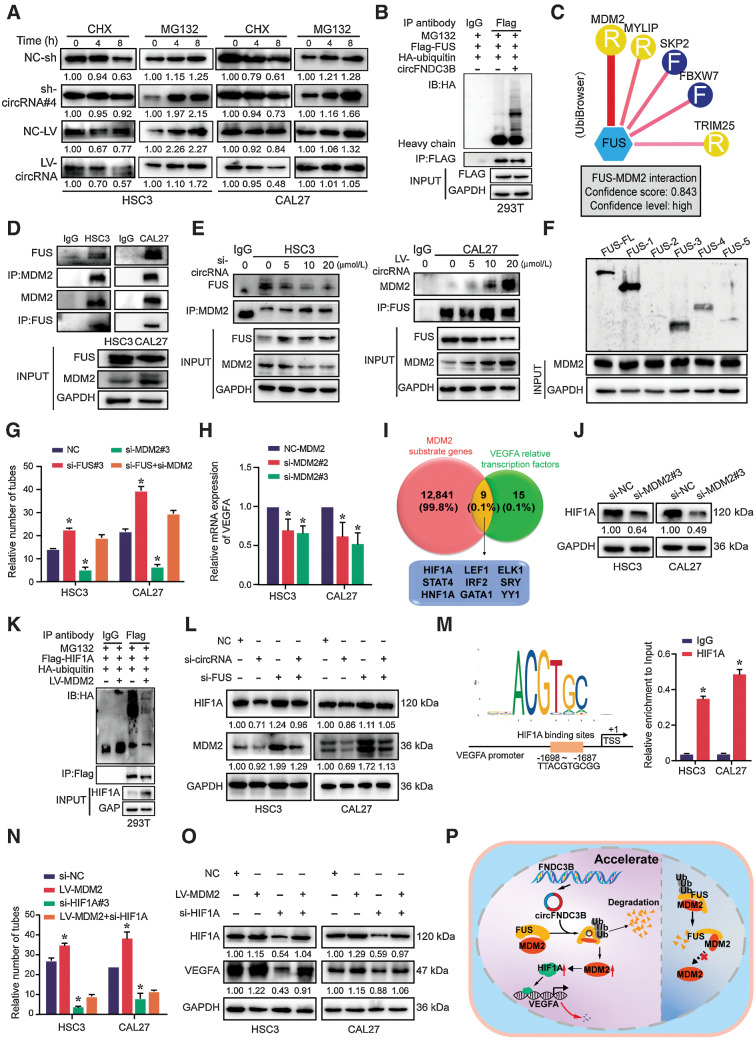
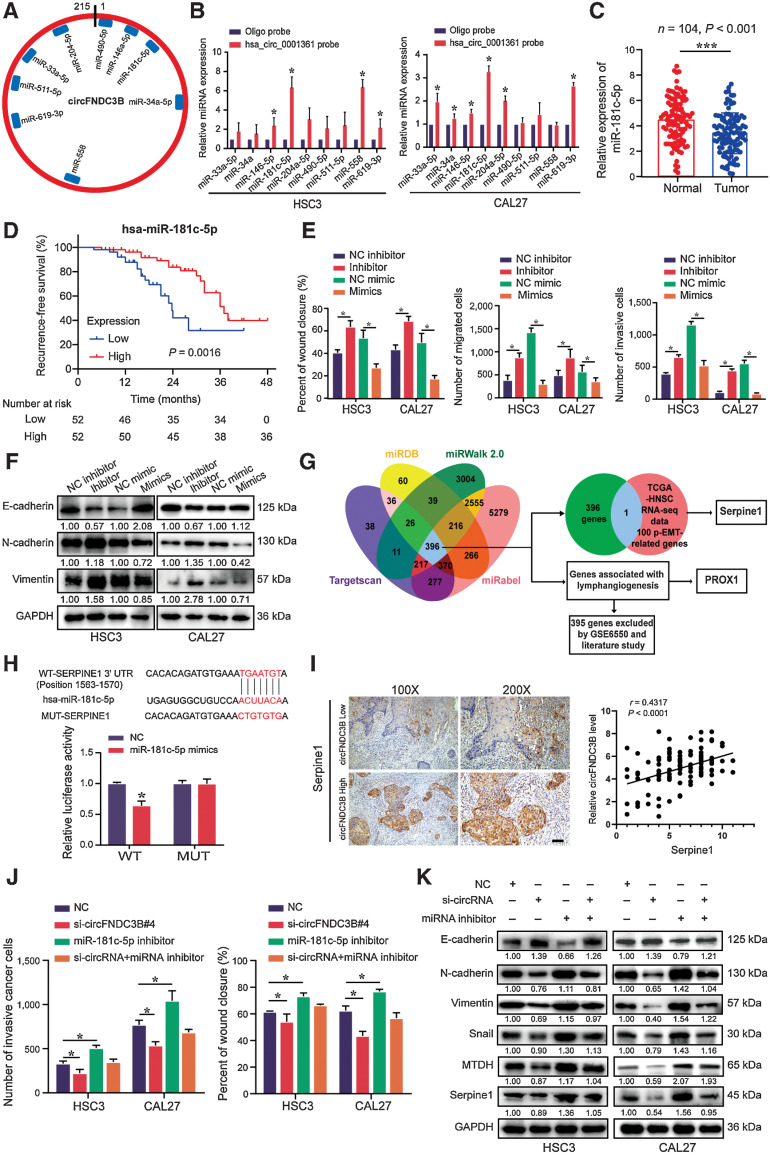
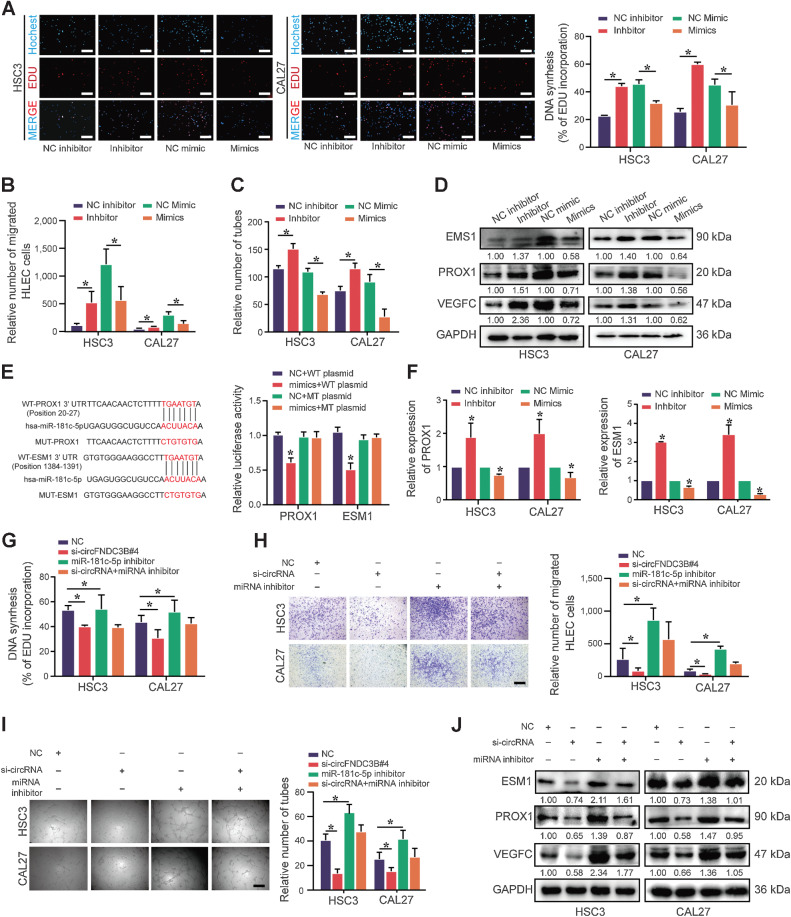
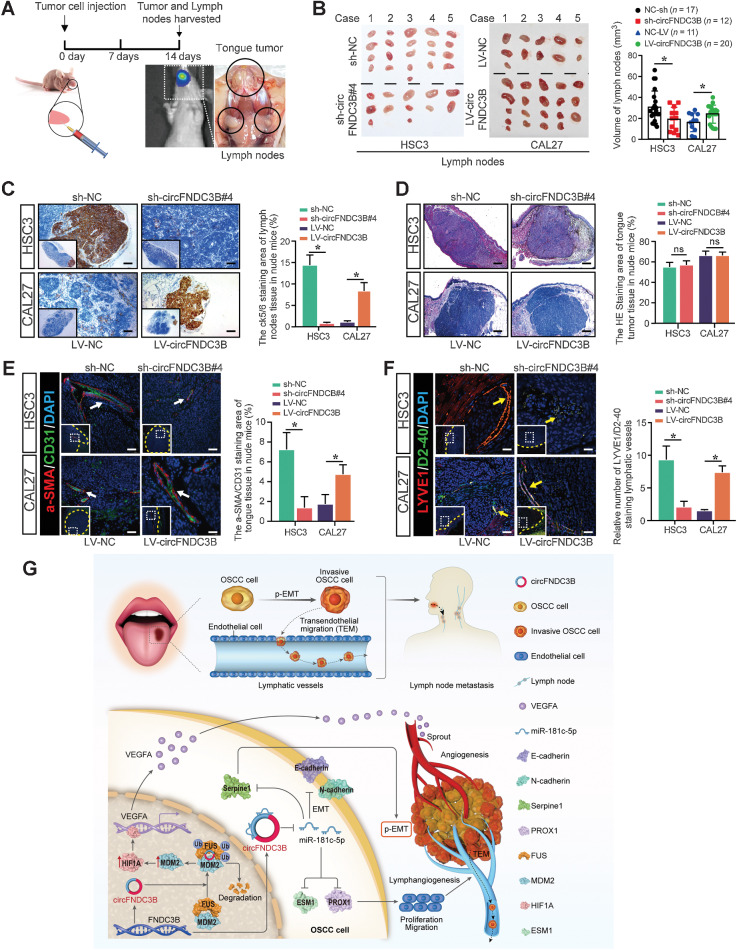
Emerging evidence has demonstrated that circular RNAs (circRNA) are involved in cancer metastasis. Further elucidation of the role of circRNAs in oral squamous cell carcinoma (OSCC) could provide insights into mechanisms driving metastasis and potential therapeutic targets. Here, we identify a circRNA, circFNDC3B, that is significantly upregulated in OSCC and is positively associated with lymph node (LN) metastasis. In vitro and in vivo functional assays showed that circFNDC3B accelerated the migration and invasion of OSCC cells and the tube-forming capacity of human umbilical vein endothelial cells and human lymphatic endothelial cells. Mechanistically, circFNDC3B regulated ubiquitylation of the RNA-binding protein FUS and the deubiquitylation of HIF1A through the E3 ligase MDM2 to promote VEGFA transcription, thereby enhancing angiogenesis. Meanwhile, circFNDC3B sequestered miR-181c-5p to upregulate SERPINE1 and PROX1, which drove epithelial-mesenchymal transition (EMT) or partial-EMT (p-EMT) in OSCC cells and promoted lymphangiogenesis to accelerate LN metastasis. Overall, these findings uncovered the mechanistic role of circFNDC3B in orchestrating cancer cell metastatic properties and vasculature formation, suggesting circFNDC3B could be a potential target to reduce OSCC metastasis.
Significance: Dual functions of circFNDC3B in enhancing the metastatic ability of cancer cells and promoting vasculature formation through regulation of multiple pro-oncogenic signaling pathways drive lymph node metastasis of OSCC.
©2023 The Authors; Published by the American Association for Cancer Research.
Figures
References
-
- Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, et al. . Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin 2021;71:209–49. - PubMed
-
- Liu J, Jiang X, Zou A, Mai Z, Huang Z, Sun L, et al. . circIGHG-induced epithelial-to-mesenchymal transition promotes oral squamous cell carcinoma progression via miR-142–5p/IGF2BP3 signaling. Cancer Res 2021;81:344–55. - PubMed
-
- SH J, Hysi D. Methods and risk of bias in molecular marker prognosis studies in oral squamous cell carcinoma. Oral Dis 2018;24:115–19. - PubMed
-
- Dieterich LC, Tacconi C, Ducoli L, Detmar M. Lymphatic vessels in cancer. Physiol Rev 2022;102:1837–79. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
